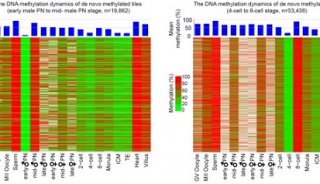
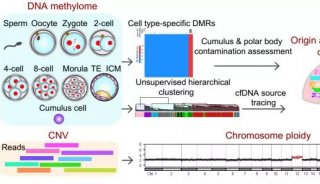
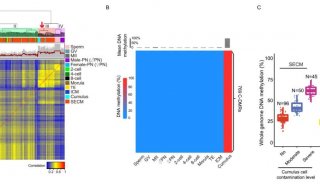
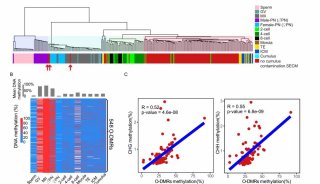
大脑中非常规的表观遗传现象
人类胚胎干细胞中有非CG甲基化,而近期研究说明在哺乳动物大脑,神经元中的非CG甲基化明显高于非神经元的脑细胞-神经胶质细胞。非CG甲基化主要发生于CA二核苷酸,额叶皮质中超过70%的非CG甲基化为mCA。有趣的是,mCH在新生的小鼠或人的大脑中缺失或只有很低的水平,但是随着出生到青春期的过渡,其含量逐渐累积。mCH的累积会短暂地与突触的发育相一致,此时逐渐成熟的额叶皮质中神经元细胞间的联系正逐渐增多。
Gabel和Chen等人提出了一种可能的机制,甲基化的CpG结合蛋白2(MeCP2)被破坏能导致神经系统疾病Rett综合征,而mCH可能就是通过对MeCP2的识别调控神经元基因表达的。Gabel等人观察到人或者小鼠中被MeCP2抑制的基因倾向于有更长的序列(核苷酸的数量),并且与mCA的数量增多相关。另一个独立的研究也支持了这一发现,该研究表明,在各种不同的神经细胞中,MeCP2的破坏更容易影响较长的基因。既往研究证实,MeCP2能与mCH相结合。Gabel等人的体外实验中进行了更为精细的分析,发现MeCP2倾向于以mCAC作为结合靶点,较之mCG,mCAC有相近或更高的亲和力。Gabel和Chen等人进行了一系列的相关分析,逐步建立了在成年哺乳动物大脑中,mCA靶向基因MeCP2结合位点的模型(如图)。Chen等人使用带有标记的表达MeCP2的转基因小鼠及染色质免疫共沉淀法构建了小鼠基因组中MeCP2结合位点的较为全面的分析图。值得注意的是,研究者发现当MeCP2被破坏后,mCH以及MeCP2结合位点区域同时富含上调和下调的基因,说明MeCP2可能有更多样的调控功能。Gabel等人的研究也支持了mCA靶向MeCP2这一模型,他们发现通过清除神经系统中DNA从头合成甲基化转移酶3a,能消除mCH的累积,进而消除对长基因的抑制,类似于破坏MeCP2的作用。
脑甲基化。当前的模型聚焦在基因组上离散的调控因子上的mCG,它可能阻止了与转录因子或MeCP2的结合,进而抑制转录。这种模型没有指出新生个体和成熟个体大脑表观遗传的差异。改进的模型强调了自出生延续至整个青春期的mCH的累积,使成熟个体的大脑呈现mCA和MeCP2高亲和力相互作用的独特现象。通过非CG甲基化对于MeCP2的募集(图中“?”表示可能的其他mCH识别因子)抑制了富含mCA的基因的表达,可能是影响了转录延长因子RNA多聚酶2。
这两个研究所揭示的机制提供了一个新的视野,让人们重新从空间和广度上认识了DNA甲基化的调控,以及靶向表观遗传学的调节机制。许多先前的研究着眼于基因表达的变化与独立调控的DNA甲基化模式的关系(主要为mCG),如启动子、CpG岛,或远处的增强子。与之相反,MeCP2优先与mCA结合,置于基因特定位点(该基因被RNA聚合酶转录的区域),并可跨越数百个千碱基对,这将有可能在个别基因水平进行精细化的基因表达调控。目前基于大规模表观遗传学分析得出的mCG局部碱基缺失与转录因子结合关系的流行观点是,启动或靶向表观遗传的调控可能与序列特异性的转录因子相关。尽管这可能是正确的,但其普适性已经被MeCP2在较长的基因上选择性靶向mCA所挑战。这是源于基因组的空间特性,并且不太可能与特异性顺式原件相关。特别的,MeCP2的募集可能是直接由表观遗传标志mCA所介导的。
mCA与MeCP2的结合形式及其涉及面目前只在全组织(如大脑皮质)或混合的神经细胞中进行研究,而将其延伸至特定的神经细胞亚型对于进一步发现mCA的模式至关重要。例如,清除MeCP2导致细胞特异性的基因调节异常。这样的实验将可以检测是否细胞特异性MeCP2调节与mCA在不同种类的脑细胞中差异性沉积相关。mCA(相比其它可能的标志物,如mCG)对于MeCP2募集还需要进一步量化,并可通过分析MeCP2结合条件性减少mCH的特定神经细胞来完成。如果mCA就是靶向MeCP2和/或其它效应因子的主要标志,那么mCH靶向长基因,并被排除在活跃表达的神经细胞基因之外的机制是什么?尽管没有对于前一个问题有说服力的解释,但对于后一个问题,答案可能是甲基转移酶3a被排除在延伸的RNA聚合酶II或与转录延伸相关的特定组蛋白修饰之外。一个相反的机制近期被发现——DNA甲基转移酶3b与组蛋白标志H3K36me3相作用,这是转录延长进而导致mC优先沉积于胚胎干细胞和原生殖细胞中活跃转录的基因上的典型标志。
推测这种表观遗传机制在正常脑功能或神经系统疾病,如Rett综合征方面的重要作用是非常诱人的。mCH在成熟个体的大脑能提供一个对接的平台去引导各种因子的结合(如MeCP2),因而可以介导成熟个体相较幼稚个体大脑所具有的独特功能。Chen等人已经开始通过寻找成熟而非幼年小鼠大脑中脑衍生神经营养因子(Bdnf)基因座上增加结合的MeCP2来研究这一联系。与增强的MeCP2募集一致,当MeCP2被破坏后,在成熟而非幼年个体的大脑中出现对Bdnf表达的错误调节。作者假设,从出生至青少年这一过程中逐步累积的mCA可能是Rett综合征迟发的原因。
在成熟个体大脑中以及多潜能干细胞中普遍存在的mCH的累积提示,除了MeCP2,可能有还未被识别的蛋白结合mCAC和mCAG,进而介导其它调控作用。寻找富含非CG甲基化的组织细胞中别的效应因子或mCH的“识读器”将扩展我们对于这一独特的表观遗传标志的认识。我们期望通过表观遗传机制的进一步研究,能使大脑中基因调控可能的例外机制得到更为全面的展现。
-
科技前沿

-
科技前沿