







P700的氧化诱导小麦叶片围绕PSI的电子传递支路
放氧光合生物通过氧化光系统I(PSI)反应中心叶绿素P700可以抑制活性氧的产生。
P700的氧化伴随着PSI中的电子流支路(AEF-1)的出现,AEF-1对于光合线性电子流(LEF)是无效的。为了表征AEF-1,我们通过测量暗区间弛豫动力学分析比较了小麦叶片二氧化碳(CO2)同化诱导期间P700和铁氧还蛋白(Fd)的氧化还原反应。在40
Pa的环境CO 2分压和21kPa的环境O 2分压下,开启1000μmolm-2 s-1光化光,逐渐氧化P700(P700
+)并提高氧化态P700 +(vP700)的还原率和还原态Fd(vFd)的氧化率。 vFd与起始点的PSII表观光合量子产率Y(II)呈正相关;
通过CO2同化和光呼吸,线性电子传递LEF调节Fd的氧化还原转换。vP700也表现出与Y(II)呈相关,但截距为正,而非零。也就是说,除了线性电子传递LEF外,PSI中的电子传递还包括电子流支路AEF-1中的电子流。这表明P700的氧化诱导电子流支路AEF-1。我们提出了潜在AEF-I的可能机制及其在减轻氧化损伤中的生理作用。
在高光,低温/高温或干旱条件下光合作用二氧化碳(CO2)同化效率的抑制会降低PSI中电子受体(NADP+)的再生效率,并增加类囊体膜PSI中的积累电子的风险。反应中心叶绿素P700是PSI中的电子源,驱动电子从质体蓝素(PC)到铁氧还蛋白(Fd),最后传给最终电子受体NADP
+的电子传递反应。完整向日葵(Helianthus
annuus)叶片置于黑暗中,反复照射高强度短脉冲闪光使PSI电子传递反应失活。短脉冲照射导致PSI受体侧电子积累,刺激活性氧(ROS)的产生,如超氧自由基和单线态氧。相比之下,在稳态光化光(AL)下的短脉冲照射处理事先启动P700氧化,就不会导致电子的积累或PSI的失活。这些数据表明电子在PSI受体侧的积累增加了 ROS产生的风险,使PSI和CO2同化失活。
光合生物利用不同的分子机制来氧化PSI中的P700。在P700得失电子转换期间,光激发P700(P700
*)氧化和/或抑制氧化态P700(P700+)还原都会加速了P700氧化。此外,黄酮(FLV)依赖的电子流和光呼吸促进P700
*氧化,以维持P700处于氧化状态。另外还有,CO2同化和光呼吸过程中的光合线性电子流(LEF)诱导类囊体膜腔内质子(H
+)的积累。腔侧的酸化抑制了细胞色素b6 /
f-复合物催化的质体醌氧化,这也有助于P700的维持氧化态。在CO2同化和光呼吸过程中,ATP合成酶介导的类囊体膜上ADP和Pi的利用控制了H+的积累。这些导致P700氧化的分子机制统称为P700氧化系统。
如上所述,黄酮FLV依赖性电子流和光呼吸中增强的电子通量有助于PSI中P700的氧化。另一方面,我们观察到P700氧化伴随着PSI中的过量电子流,这不完全是由LEF驱动的;这种过量的电子流被称为PSI中的替代电子流(AEF-1)。在这项研究中,我们使用DUAL /KLAS-NIR分光光度计(Walz,德国)对小麦(Triticum aestivum)叶子中的AEF-I进行了分子表征,这是一种新型分光光度计,专门检测P700+,氧化态PC(PC+)及还原态Fd(Fd-)。在诱导AEF-1期间,我们评估了P700,PC和Fd的氧化还原状态之间的关系。此外,我们使用暗区间弛豫动力学(DIRK)分析研究了P700+和PC+的还原速率与Fd-的氧化速率之间的关系。这些分析帮助我们了解调节AEF-1活化的机制以及P700氧化与AEF-1活性之间的关系。此外,我们研究了光合生物中P700氧化的生理功能。 P700的氧化还原周转率远高于Fd,并且显示出对小麦叶片中LEF的电子通量的绝对依赖性。换句话说,AEF-1中的电子通量由P700氧化诱导并在PSI内起作用。另一方面,P700氧化有助于Fd-的氧化,从而揭示了P700氧化的新功能。我们通过抑制PSI中ROS的产生,提出了AEF-1的分子机制及其在减轻氧化应激中的生理功能。

DIRK分析关闭光化光(AL)后小麦叶片中氧化态P700(P700 +),PC(PC+)和还原态Fd(Fd-)的衰减。使用Dual / KLAS-NIR分光光度计实时监测P700,PC和Fd的氧化还原状态的变化。为了确定在光化光条件下小麦叶片中P700+和PC+的还原速率和Fd-的氧化速率,在坐标时间0点瞬时关闭AL并延续测量400ms。在坐标时间0时刻P700+,PC +和Fd-的下降的初始斜率表明P700+和PC+的还原速率和Fd-的氧化速率。P700+,PC+和Fd-的初始斜率变化的特征是在叶温25°C,O2分压21 kPa 和光强1000μmolm-2s-1下测量70组平均得到的,A :CO2分压40 Pa,B:CO2分压5Pa ,AL关闭后持续110 ms。这些数据是在稳定状态下获得的,通过稳定Y(II)的测量得到证实。
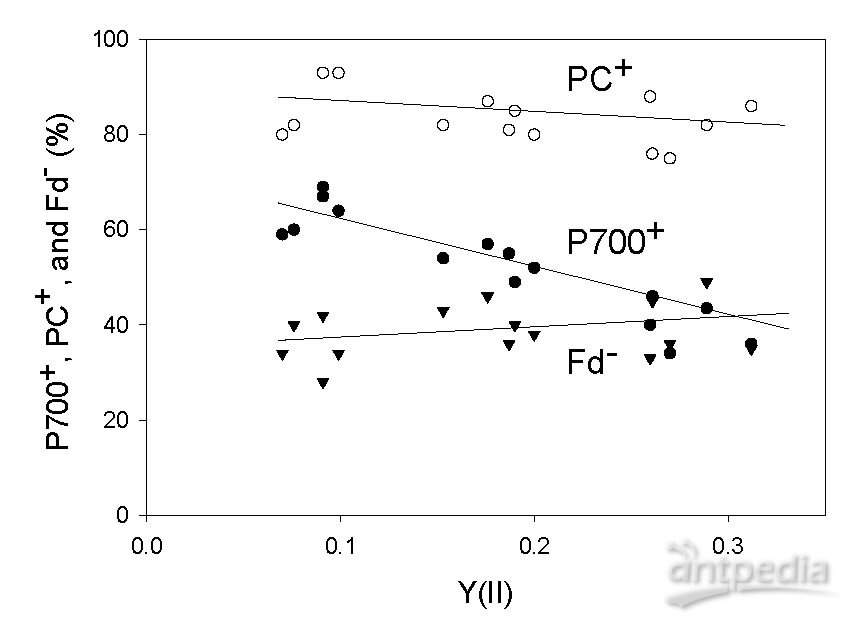
小麦叶片P700 +,PC +和Fd-与表观Y(II)的关系。 P700 +,PC +和Fd-及表观Y(II)均在叶温25°C,O2分压21kPa 和光强1000μmolm-2 s-1下测量。CO2分压从40Pa到20Pa再到5 Pa逐步降低。 P700 +,PC +和Fd-的数据来自5个平行植株。 CO2分压的下降使得Y(II)下降。通过获得稳定的Y(II)证实了在几个CO2分压下测量时达到稳态的。空心圆,PC+;实心圆,P700+,倒三角形,Fd-。

小麦叶片中vPC,vP700和vFd与表观Y(II)的关系。数据点表示为光化光关闭后5ms的相对变化(%),vPC,vP700和vFd与表观Y(II)的数据来自5个平行植株光化光关闭后的初始变化,CO2分压的下降使得Y(II)下降。空心圆,vPC;实心圆,vP700; 倒三角形,vFd。

小麦叶片光合诱导的P700+、PC+和Fd-的响应。在0 s处,打开光化光AL照射小麦叶片。在叶温25°C,O2分压21kPa ,CO2分压40Pa和光强1000μmolm-2 s-1下测量P700+(A)、PC+(B)和Fd-(C)。P700+、PC+和Fd-的响应数据来自5个平行植株。I、时期I;II、时期II;III、时期III。

小麦叶片光合诱导的vP700、vPC和vFd的响应。在0 s处,打开光化光AL照射小麦叶片。在叶温25°C,O2分压21kPa ,CO2分压40Pa和光强1000μmolm-2 s-1下测量vP700(A)、vPC(B)和vFd(C)。vP700、vPC和vFd的响应数据来自5个平行植株。在指定的时间点引入DIRK分析的瞬态(400ms)黑暗(关光化光)。数据为mean±SD(SD;n=5)。I、时期I;II、时期II;III、时期III。

AEF-1的假设途径。光激发P700(P700 *)向第一个电子载体A0提供电子,产生P700 +。随后,P700+接受由PC传递的来自PSII的电子以完成P700再生。A0将电子提供给第二电子载体A1。此后,电子分别通过第三和第四电子载体Fx和FA / FB流向铁氧还蛋白(Fd)。空心箭头表示光下氧化还原循环中的P700周转。实线箭头表示电子流动。虚线箭头表示电荷重组过程中的电子流动。电荷复合是附加电子流动的机制之一。
