等电聚焦和二维凝胶电泳实验
对于复杂混合物,如全细胞裂解物或富集的亚细胞组分,通过两步正交的分离 (orthogonalseperation),二维凝胶 (2D 胶)电泳可以很好地分离成几百个至上千个单个蛋白质。第一次分离基于电荷,即使用变性等电聚焦电泳; 第二次分离基于表观分子质量,即使用变性十二烷基磺酸钠-聚丙烯酰胺凝胶电泳 (SDS~PAGE)。二维凝胶实验在观察经带电荷的翻译后修饰的蛋白质的亚型方面效果极好,如磷酸化作用和硫酸化作用 (增加电荷)、或乙酰化作用(中和电荷)。
| 试剂、试剂盒 | |
|---|---|
| 仪器、耗材 | |
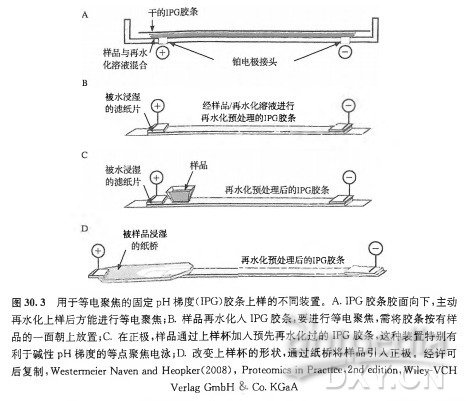
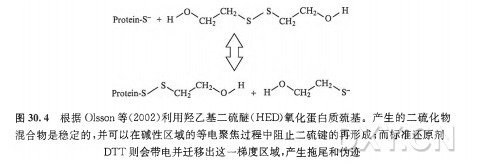
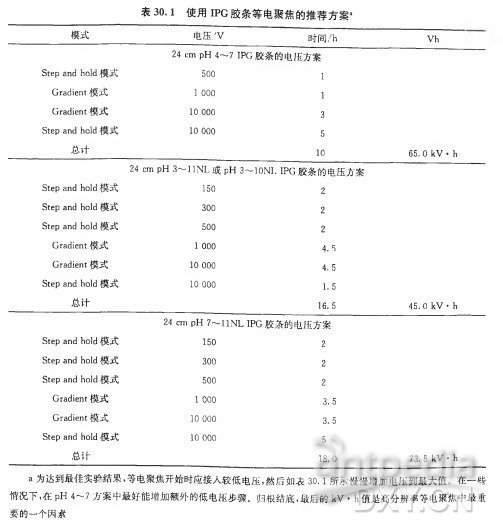
| 实验步骤 | 一、等电聚焦的基本原理等电聚焦 (IEF) 是一种能根据分子内的质子接受点的 pKa 值所决定的电荷量分离两性分子 (如蛋白质和肽类) 的电泳分离方法。对于蛋白质和肽类而言,这些点位可以位于 N 端和 C 端的自由氨基及羧酸,也可以位于精氨酸、赖氨酸、组氨酸、天冬氨酸和谷氨酸残基的侧链。等电点是两性分子特有的物化参数,它是分子净电荷为零时的 pH。等电聚焦具有极高的分辨能力,包括分离能够改变电荷的蛋白质翻译后修饰 (如磷酸化作用、乙酰化作用/去乙酰化作用)。虽然这一方法可应用于若干类型的两性复合物的分离,但章节将只讨论蛋白质分离的方法论。 等电聚焦电泳样品的制备需要高浓度离液剂、两性离子去污剂、还原性巯基及载体两性电解质,以防止在蛋白质之间形成聚集物和复合物。制备高分辨率的等电聚焦电泳, 必须确保样品中不存在非蛋白质离子,因为需要保证高电阻以在最小电流的状态下产生高电场 (8000~10000V)。通常用蛋白质沉淀法减少此种污染物 (见本章 3.2 节)。理想情况下,等电聚焦电泳样品中的唯一离子便是蛋白质本身。随后,该样品便用于含有与样品相同添加物的等电聚焦凝胶中,并在电场中分离。在蛋白质到达其等电点后,将凝胶孵育在 SDS 缓冲液中,再根据蛋白质表观分子质量,用 SDS-PAGE 平板凝胶进行分离(分子质量是主要影响因素, 但也受到疏水性影响, 并且蛋白质形状也有着较小影响)。 用于二维凝胶电泳的现代等电聚焦法采用薄聚丙烯酰胺凝胶作为分子筛,该分子筛具有固定的 pH 梯度(IPG)。这种凝胶中的丙烯酰胺浓度较低(丙烯酰胺总量通常为 4%~5%), 因为此基质不能对高分子质量蛋白质有所限制。将蛋白质加入此基质中,再加上电场。在 pH 梯度上低于 (也就是酸性强于)自身 pi 的蛋白质将带正电荷,并向电场的阴极移动。相反地,在 pH 梯度上碱性强于自身 pi 的蛋白质将向电场的阳极移动。Pl 表示蛋白质电中性时的 pH,因此当蛋白质在 pH 梯度上的值等于 pi 时,不受电场影响。 这样一来,蛋白质便会停止移动,并且牢牢保持在与自身 Pl 相等的 pH 处。这一方法具有得天独厚的聚焦效果—当蛋白质从其 Pl 点处扩散时,便会带上电荷,从而电场又会迫使其移回等电点。 在等电聚焦过程的最后,蛋白质便会高度聚集在其等电点上,因此探测时灵敏度非常高。即使是微小的电荷差异也能区别开,并且可以采用更大的分离距离和更小的梯度间隔来提高分辨率 (HovingetaL,2000)。如有需要,还可以用蛋白质标准品标定校准曲线来估计蛋白质的等电点。 等电聚焦的 pH 梯度类型1.载体两性电解质在由自由溶液中的混合两性缓冲液构成的 pH 梯度中分离蛋白质这一概念,是由 Vesterberg 和 Svensson(1966) 提出的,Vesterberg 在几年后便将这一概念转化为可行的方法 (VeSterberg,1972)。由于自然形成的氨基酸和多肽在其等电点时具有很弱的缓冲能力,所以缓冲液需要通过化学方法进行合成。 Ampholines(两性电解质) 是首批以此为目的而投入市场的试剂,该试剂是脂肪族寡氨基酸和寡碳酸的混合物,包含 600~700 种不同的同系物,这些同系物的等电点为 pH3~10。这些复合物在等电点时具有很强的缓冲能力,并能在电场的作用下形成 pH 梯度。混合物的分子质量小于 IkDa 时最适宜,并且因为高亲水性不会与蛋白质结合。 等电点范围明确后,将有助于提高 pH 梯度相差较小的混合物的分辨率。 载体两性电解质是近年发展起来的,与 Amphonlines 试剂相比,它们是由不同种的试剂合成的,并可从多种商业渠道中购得。尽管这些产品都能在电场作用下形成 pH 梯度,但它们的 pH 梯度具体分布、缓冲力的分布以及同系物数量却是不同的(Righettietal.,2007), 因此它们的聚焦模式也不尽相同。 基于载体两性电解质的等电聚焦大多是在聚丙烯酰胺凝胶中进行的,O’Farrell(1975) 提出的最初的二维凝胶电冰过程在薄凝胶管 (一维时为柱状胶) 中的载体两性电解质梯度中完成等电聚焦。但是这些长软型的凝胶不易操作,且随着电泳时间增加,pH 梯度也会变得不稳定, 从而引起阴极漂移。虽然一些实验室仍然采用载体两性电解质技术,但是这一技术方法大多被 IPG 胶条所替代,我们将在下面讨论 IPG 胶条。 2.固定的 pH 梯度随着用于等电聚焦的聚丙烯酰胺基质中固定 pH 梯度这一概念的引人,二维凝胶电泳在方法论上有了重要的发展(Bjellqvistetal.,1982)。这一重要的技术发展克服了载体两性电解质系统的一些不足, 如梯度漂移 (特别是在阴极区)、机械不稳定性以及不同次电泳之间和实验室之间的技术差异。IPG 胶条技术极大地推动了二维电泳方法论, 并大大提高了实验室内部及不同工作组间的重复性 (Gorgaal.,2000;2004),如今已经成为等电聚焦的备选方法,并可从一些商业渠道获得。 在现代 IPG 胶条技术中,pH 梯度是由酸性和碱性缓冲组组成的。这些缓冲组是在制备凝胶时与聚丙烯酰胺基质共聚合而成的。只需要不到 10 种不同的具酸性和碱性 PKa 值的丙烯酰胺衍生物便足以获得任意的 pH 梯度。需要制备 2 种含有计算好的丙烯酰胺衍生物混合体的单体溶液: 酸性梯度端溶液的制备需加入一部分甘油用以在倒胶时稳定梯度。这些 PH 梯度凝胶的铺制是以共价结合的薄膜衬底为支撑的。在凝胶制备完成后,需用蒸馏水将聚合催化剂和非反应混合物从介质中洗去,以便产生等电聚集所必需的极低电导 (Westermeier,2005)。 IPG 胶长期保存时需干燥,并需在使用前不久进行再水化处理。IPG 胶的主要好处在于不存在阴极漂移问题,因为梯度是固定于基质上的。IPG 胶主要应用于变性条件下的等电聚焦,作为高分辨率二维凝胶电泳的第一维所使用; 其中仍然使用载体两性电解质,并且为了提高导电性和蛋白质的溶解度,在样品中和 1PG 胶的再水化溶液中都要加人载体两性电解质。 如今可以从一些供应商处购买到预制的 IPG 胶条,这些胶条具有各种长度和 PH 梯度,可以实现蛋白质裂解产物中选定的蛋白质组分的最优化分离与呈现。尤其是,胶条的 pH 梯度有非常宽的范围 (如 pH3~11),有中间范围 (如 pH4~7、pH7~11),还有狭窄范围 (如 pH4~5、pH5.5~6.5)。胶条长度同样非常多样,短至 7 cm, 长至 24 cm。典型胶条宽度为 3 mm,平均厚度约为 0.5 mm(图 30.2)。为了实现预期实验目的, 需要挑选具有最优 pH 范围和长度的胶条 (Hovingetal.,2000)。例如,7 cm、pH3~11 的 IPG 胶条可以为小样式凝胶提供最好的范围,但是总体来说,它们为蛋白质组范畴的、基于探索性质的实验提供的分辨率和灵敏性是最低的。 IPG 胶条提供几种样品上样模式。再水化上样和杯状上样是两种主要使用的方法,还有主动再水化的修改方法和纸桥上样(图 30.3)。本章 3.3 节和 3.4 节对这些方法有非常具体的介绍。样品上样方法的选择取决于 pH 梯度的类型和样品。因为 IPG 胶条有一个平整的表面, 所以如果需要, 可以应用于水平平板 SDS 凝胶电泳。 SDS-PAGELaemmli(1970) 描述的 SDS-PAGE 在很长一段时间内作为各种生化分析中分辨完整蛋白质的备选方法。这主要是因为对于疏水性很强的蛋白质,SDS 是最好的增溶去污剂,所有的蛋白质,包括碱性很强的蛋白质,都向同一方向移动, 分离取决于各自的表观分子质量 (通常称为分子质量)。 1.分离原理当将过量的 SDS 加入到蛋白质溶液中时,蛋白质会形成一些阴离子微粒,其每个质量单位带一个恒定的净负电荷。蛋白质的三级结构和二级结构被破坏,肽链被解开。为了使肽链完全展开,需要使用诸如 2_巯基乙醇或二硫苏糖醇等还原剂把半胱氨酸之间的二硫键打开。Ibel 等 (1990) 认为, 这些复合物的多肽表面部分被覆盖,部分被打开,犹如项链一般。在电泳过程中所有的 SDS 微粒都移向正极, 其电泳迁移率主要取决于分子质量,但也会受蛋白质疏水性的影响。如果将蛋白质的迁移距离对表观分子质量的对数作图,就会得到一条在线性范围内的 S 形曲线。在共迁移的蛋白质分子质量标准的辅助下,就可以估算多肽的分子质量。 2.胶的类型一般来讲,SD&PAGE 可以在各种各样的基础缓冲体系中进行;但是,LaemmIi(1970) 创立的不连续缓冲体系是最常用的体系, 在胶中使用 PH8.8 的 Tris-HCl, 在电泳缓冲液使用 SDS^Tris-甘氨酸。对于一维分离,pH6.8 的低浓度积层胶使样品缓慢进入胶内,而不发生聚集,而且在分离起始时有压缩条带的效果。但是, 在二维凝胶电泳中通常不使用积层胶,因为蛋白质已被预分离,而且已经在 3 mm 宽的 IPG 胶条内被高效浓缩 (详见本章 3.6 节)。 标准的基质是含有 12.5%T(全部为丙稀酰胺) 和 2.6%C(用于交联的双丙烯酰胺) 的均一胶层。但一些应用中对于确定的范围需要精确分离。例如,对于高分子质量蛋白质需要更低的%T; 而对于低分子质量蛋白质,更高的会有更好的解析度, 所以浓度要提高。梯度 SDS-聚丙烯酰胺凝胶可以提供更大动态范围的分辨率, 但是其灌胶的一致性和可重复性是极大的挑战,但这些要求对于使用二维凝胶的定量蛋白质组学是至关重要的。多孔梯度凝胶显示出更宽的分子质量线性范围, 并且可提高一些较难解析的蛋白质的分辨率, 如糖蛋白。 SDS 电泳可在夹在玻璃板中的盒式垂直装置中运行,也可在水平平面上进行。胶的厚度为 1.5~ 0.5 min。较厚的凝胶在力学上更加稳定,但是更难染色,而且会产生额外的样品损失,对随后的质谱分析也会产生额外的背景噪声。较薄的凝胶更易有效地冷却,因此电泳可以进行得更快从而有更好的分辨率。 相对于过去几十年中二维凝胶流程取得的进展,二维 SD&PAGE 分离几乎没有什么变化。垂直电泳系统的使用更普遍,同时跑 1 块或 12 块甚至更多块胶的设备也已经出现。在历史上水平平板装置也是可行的, 而且因其低试剂消耗量 (如 SDS 电泳缓冲液)、样品的处理方法以及较短电泳时间带来的提高分辨率的潜力,现在有复兴的迹象。一些进展包括: 使用有弹性的塑料支持物 [包括用于差异凝胶电泳 (DIGE) 和其他基于荧光的技术的弱荧光介质], 用于同时灌制多块可重复的丙烯酰胺梯度和不同丙烯酰胺配方的凝胶的特别装置,以增加凝胶的保质期和耐用性。 碱性 pH 梯度分辨率的提高相对于酸性 pH 梯度, 碱性 pH 梯度的高分辨率等点聚焦更具有挑战性,这既是由于阴极漂移的作用,又是由于维持蛋白质可溶性的还原剂的丢失(DTT 是弱酸, 在 pH 大于 8 时带电,因此会与梯度的碱性区域分离)。阴极漂移作用最早通过引人非平衡 pH 梯度电泳 (NEpHGE) 得到解决 (OFarrelletal.,1977)。但是在 pH 大于 8 时还原剂丢失的问题仍然存在,而且由于半胱氨酸不再受到保护,导致多肽与尿素发生非特异反应、肽段背向折叠 (backfolding) 和聚集,造成二维凝胶碱性区域中蛋白质点的伪迹 (artifact) 和水平拖尾(streaking)。 用膦类化合物,如 TBP 或 TCEP 替换巯基试剂可以部分防止这类效应,但是由于这些试剂在电场中不稳定,会产生额外的伪迹。在等电聚焦之前,用碘乙酰胺、乙烯基吡啶或单体丙烯酰胺预先将蛋白质烷基化,都会因多肽烷基化不完全及等电点的变化而产生额外的伪迹。 过量使用羟乙基二硫醚(hydroxyethyldisulphide,HED)[商品化后商品名为 DeStreak(GEHealthcare)] 提供了 pH 下保持疏基处于还原状态(通过质量作用)的方法,显著提高了使用 IPG 胶条的碱性等点聚焦电泳的分辨率 (Olssonetal.,2002)。特别是在阳极样品人口联合使用上样杯上样或纸桥上样时效果更佳,因为上样时碱性蛋白质都带着相同的酸性电荷进入梯度胶条,然后向阴极移动从而达到等电点 (而不是令它们向两个方向都移动)。 当蛋白质进入到 IPG 胶条时,半胱氨酸侧链上的巯基会立即被氧化成高度稳定的二硫化物混合物。这是一种高特异性的,可以阻止不需要的副反应的平衡反应(图 30.4)。 使用 DeStreak 方法制备样品时可以采用小量的 DTT,那么蛋白质便可在聚焦前保持还原状态。然而需要注意的是,过量的 DTT 会将 HED 还原成 2-巯基乙醇, 这会导致更多拖尾①。 差异凝肢电泳即便 IPG 胶条重复性越来越高,样品制备和电泳条件更加严格,凝胶之间还是存在差异。差异表达蛋白质组实验的成功依靠对单独采集 (生物学上的) 样本的重复的测量,同样也需要技术上的重复 (对同一生物样本的重复测量)以控制分析中产生的差异,如样品采集、处理和凝胶之间的变化。通过运用 DIGE 技术 (FriedmanandLilley,2009;LilleyandFriedman,2004;Unluetal.,1997) 可以解决这些方面的挑战。 1.染色和检测近来,二维凝胶电泳所用的荧光染料在发展和应用上获得了重大进步。这些染料用于检测和定量溶解在凝胶中的完整蛋白质种类。荧光染料特别提供了检测的敏感性,其敏感程度至少大于等于银染法(约 Ing),并且还将丰度的线性动态范围极大地提高了 3~4 个数量级 (银染和考马斯亮蓝 R-250 染色法通常提供低于 1 个数量级的动态范围)。 2.DIGE 和定量分析算法1997 年,DIGE 法开始使用 (Unluetal.,1997),此法将混合样品的荧光标记的动态范围与灵敏度应用于同一块胶中,排除了共分析样本中的分析性偏差 (几种凝胶之间的)。 现在这一方法也能在一系列 DIGE 胶中应用,利用混合的样本内对照,使迁移模式配准成为可能,从而标准化丰度比(abundanceratio),为多变量实验提供了非凡的统计支撑 (KarpandLilley,2005)。 简而言之,这一方法要求事先用光谱区别明显的荧光染料 (Cy2、Cy3 和 Cy5) 标记多种样本,然后将样本混合并在同一二维凝胶中分析。这一方法能排除几种凝胶之间的偏差,并且相互独立的突光影像也能被单独记录,用于单个分辨特征 (resolvedfeature) 中的蛋白质丰度变化的直接定量(图 30.5)。这里用到了两种不同的标记概念:「最少量」标记,即将占蛋白质总量约 5% 的赖氨酸中的 e-氨基进行标记广充分」标记,即将蛋白质混合物中所有可标记的半胱氨酸「充分饱和」地进行标记。 在一系列含有不同的由 Cy3 或 Cy5 标记的凝胶中分析用 Cy2 标记的内参照时,DIGE 法最见成效,并有统计依据。重要的一点是,该 Cy2 标记的内参照是由实验中所有样本等量混合而成,并在多凝胶实验的每一块凝胶中都存在。由于个体样本(由 Cy3 或 Cy5 标记) 与等份的 Cy2 基准混合物相混合,每种分辨特征 (resolvedfeature) 便可直接与凝胶中 Cy2 基准混合物中的同源特性(cognatefeature) 联系起来。凝胶中的 Cy3:Cy2 与 Cy5:Cy2 之间的各种比例便能在不受几种凝胶之间偏差的干扰之下算得,这些比例可用于实验中另一些样本中这一特性的所有其他衡量标准,不但技术性 (分析性) 误差极小,而且有着强有力的统计学功效 (KarpandLilley,2005)。 DIGE 法也直接适用于多变量统计分析之中,如主成分分析和系统聚类。这些附加的统计工具能够有效地帮助人们发现一组实验样本中的差异。重要的是,还能帮助确定差异中的主因素是否能够描述生物性,或者能够揭示样本之间未曾预料到的 (或是在准备样本时被带入的) 差异,或者能够精准定位出能对实验刺激或分类作出集体反应的蛋白质亚群 [如见 Franco 等(2009);Friedman 等(2006;2007);Hatakeyama 等(2006);Suehara 等 (2006)。综述见 Friedman 和 Lilley(2009);Lilley 和 Friedman(2004)]。 3.软件分析工具现在,一些软件程序能辅助完成二维凝胶实验 (无论是 DIGE 还是其他) 的定量分析。 这些程序最大的区别,便是在探测蛋白质特性 (界线)和几种凝胶之间校准(aliSnment)/配准 (registration)(如基于载体的图像变形)时的算法有所不同。总体而言,这些程序都提供了强有力的伴有单变量 (如 f 分布检验和方差分析) 和多变量 (如主成分分析和系统聚类)的统计分析的分析工具, 非常利于估量个体蛋白质特性和整体表达模式的丰度变化,该类模式能够将描述生物学表型的变化和那些在实验中未预料的偏差辨别区分。 二、方法蛋白质样品制备稳定的样品制备对于任何成功的生物分析性测定都是至关重要的。为了增加实验的重复性, 并将预期外的变异降至最小,使用的缓冲液和材料都应该是质量最好的,并且在采购时需特别小心。应该使用小分子蛋白酶和磷酸酶抑制剂,如抑肽酶 (aprotinin)、亮抑肽酶 (Ieupeptin)、胃蛋白酶抑制剂 (pepstatinA)、木瓜蛋白酶抑制剂 (antipain)、4-(2-氨乙基) 苯磺酰氟 (AEBSF)、铒酸钠(sodiumorthovanadate)、冈田酸(okadaicacid) 及微囊藻素 (microcystin) 等 (或采用这些小分子抑制剂的商品化试剂盒)。值得注意的是,不要采用会在二维凝胶中溶解的抑制剂 (如大豆胰蛋白酶抑制剂)①。 (1) 只要样品随后会沉淀以移除非蛋白质离子组分 (可严重干扰等电聚焦的成分, 详见本章 3.2 节), 则基本上可以使用任何一种蛋白质提取缓冲液。随后用一种与二维凝胶相兼容的缓冲液重悬样品。在许多情况下蛋白质提取物无需沉淀,直接在这种缓冲液中制备和分析。不用将额外的离子化合物加入到样品中,下述缓冲液已含有充分的高离液活^性(chaotropicactivity)。 ①标准二维凝胶电泳裂解缓冲液 (RabiUoucUl 卯 8):7mol/L 尿素、2mol/L 硫脲、4%CHAPS、2 mg/mLDTT 及 50 mmoVLpH8.0 的 Tris-HCU ②用于膜相关蛋白质时有所调整:7mol/L 尿素、2md/L 硫脲、2% 脒基磺基甜菜喊-14 及 50 mmol/LpH8.0 的 Tris-HCl。缓冲液系统包含其他盐和去污剂,尤其是十二烷基磺酸钠,可更有效地提取蛋白质,但必须在进行等电聚焦前先沉淀。 ③TNE:50 mmol/LpH7.6 的 Tris-HC1、150 mmol/LNaCl、2 mmol/LpH8.0 的 EDTA、2 mmol/LDTT 及 l^WNP-40。 ④RIPA 缓冲液:50 mmol/LpH8.0 的 Tris-HCl、150 mmol/LNaCl,l%NP-40、0.5% 脱氧胆酸及 0.1%SDS。 (2) 迅速重悬细胞以抑制蛋白酶解活性是至关重要的。将提取物 15000 g 或 100000 g 离心 10 min,除去可干扰等电聚焦的不溶性蛋白质和磷脂。 (3) 某些情况下,超声可断裂核酸使其在随后的净化步骤中与磷脂一同被除去,从而提高样品质量。核酸和磷脂这两种非蛋白质类阴离子组分都会干扰等电聚焦 (详见本章 3.2 节)。建议将样品放置冰上,并使用超声仪尖端探头进行短脉冲处理。 (4) 在所有的步骤中,使体系保持冷环境是很重要的, 尤其是含有尿素的样品绝不能被加热。尿素过热 (超过 37°C) 会加速异氰酸盐 (一种尿素的自然分解产物)的形成,反过来将使自由氨基甲酰化。当该反应发生在蛋白质上时 (发生在氨基末端的残基上或在赖氨酸残基的 e-氨基上),会阻止这些位点的质子化,引起等电点的酸性漂移。二维凝胶电泳中大量样品氨基甲酰化造成的结果是漂亮的蛋白质斑点串,看起来好像是翻译后修饰,但却完全是伪迹。 (5) 测量蛋白质浓度可采用各种标准方法。需注意要采用与蛋白质提取缓冲液相兼容的方法。例如,CHAPS 和硫脲(尽管完全适合于蛋白质提取)将会干扰 Bradford 或 BCA 实验,导致数据错误和不可靠。遇到这些情况时,应该在定量前沉淀分装出来的小份样品,再重溶于适当的缓冲液中,或采用与定量实验相兼容的去污剂。 (6) 对于细胞培养实验,蛋白质浓度可由细胞数目估算得出。例如,对于由 697 前-B 淋巴瘤细胞株制备得到的蛋白质样品(可从德国微生物菌种保藏中心获得,DeutscheSammlungvonMikroorganismenundZellkulturenGmbH,DSMZNo:ACC12),IO7 个细胞约对应 Img 蛋白质提取物。为了移除生长培养基组分 (尤其是血清蛋白质),在收集细胞前必须要彻底地清洗细胞 (至少用 PBS 清洗 2 次)。需用细的移液器 tip 头将最终的细胞沉淀中所有的 PBS 小心吸去。细胞沉淀 (通常每管 IO8 个细胞) 保存在一 8 0°C。 (7) 细胞提取物此时可以用于脱水的梯度胶条,可以通过胶内再水化或是杯上样完成。在加人样品前,应该加人适当的两性电解质或是 IPG 缓冲液,大多数只需 0.5%(V/V)(但如果需要的话增量到 2% 也可以接受)。如果样品缓冲液中没有溴酚蓝,加入少量 (可加入干固体的少许晶粒, 或者几微升溴酚蓝水溶液) 以作为等电聚焦的失踪染料,同时也作为杯上样过程中的视觉辅助。或者,样品可无期限地先保存在一 8 0°C。 样品净化与沉淀正如前述,有时非蛋白质离子 (如盐、磷脂) 的存在会降低 IPG 胶条的电阻, 干扰等电聚焦,因此在不使胶条过热的情况下难以获得需要的高电压以进行高分辨率聚焦。由于大部分商品化的等电聚焦仪器都将许多胶条集合在并联电路上,其中一个胶条出现电阻显著性差异将会对其他胶条分离造成不利影响。 通常一 欠净化或沉淀步骤可以移除这些干扰离子,或者至少使所有样品标准化至具有相似电阻。沉淀不仅可以同时移除多种污染物,而且还能有效瓦解复合物和不可逆地抑制蛋白酶活性。最有效的沉淀方法是使用甲醇和三氯甲烷 [根据 Wessel 和 Flugge(1984)] 或使用三氯乙酸、脱氧胆酸盐和丙酮 [根据 Arnold 和 UlbriCh-H o fmann(1999)]。 在等电聚焦前,蛋白质必须在增溶液中再溶解 (详见本章 3.1 节)。此外,一些用于蛋白质二维凝胶分析的沉淀试剂盒也可从一些商业渠道获得。Wessel 和 Flugge(1984) 方法的改编版描述如下。 (1) 将预定量的蛋白质提取物用水补足体积至 1 00 pL。 (2) 加人 3 0 0 水 (3 倍体积)。 (3) 加人 400 甲醇 (4 倍体积)。 (4) 加人 100fiL 氯仿 (1 倍体积)。 (5) 剧烈涡漩振荡并离心。蛋白质沉淀物将出现在分界面上。 (6) 将水/甲醇混合物从分界面顶端去除,小心不要扰动分界面。沉淀的蛋白质通常不会形成一个可见的白色分界面,因此注意不要破坏分界面。 (7) 再次加人 400 甲醇以清洗沉淀。 (8) 剧烈涡漩振荡并离心。蛋白质沉淀物便沉积于试管底部。 (9) 去除上层清液,并用真空离心机将蛋白质沉淀简单地干燥。 (10) 在适量的可兼容二维凝胶的缓冲液中重悬这些蛋白质沉淀 (详见本章 3.1 节)。 当用沉淀法作为净化步骤时,建议蛋白质起始浓度为 1~1 0 mg/mL。如果蛋白质样本过稀,在清除沉淀后将很难定量地回收蛋白质。冻/融也应控制在最少量,通常 ImL 分装量或更少的冷冻样本就已足够。对于 DIGE 实验而言,沉淀步骤极大地满足了标记样本在不含游离氨且酸碱平衡的二维凝胶电泳样本缓冲液的需求。 使用酸性范围的 IPG 肢进行等电聚焦大多数商品化的等电聚焦仪器每次电泳时都能够装载至多 12 条独立的 IPG 胶条。 重要的一点是,要将同一次电泳中待测的每根 IPG 胶条都尽可能保持同一水平,因为在大多数配置中,单个的 IPG 胶条会在正电极和负电极之间形成一个并联电路。任何 IPG 胶条若在组分中 (特别是在离子成分方面) 异于其他胶条,便会影响到所有胶条的分辨率,有时这种消极影响是巨大的。因此,强烈建议只同时聚焦同一 PH 梯度的相同长度的 IPG 胶条,所用的样品也要来源于同一实验(如样本类型和组分要尽可能相同)①。 以下方法用于 24 cm,pH4~7 的 IPG 胶条。 (1) 将 IPG 胶条在含有蛋白质总量高达 0.5~2 mg 的二维凝胶样品缓冲液中再水化 (详见本章 3.1 节),对于每根 24 cm 的胶条,需要在再溶胀托盘中将蛋白质样品稀释或重悬至终体积 450uL。用 2~3 mL 的石蜡油覆盖胶条防止尿素结晶。 (2) 加人样品的胶条的再水化过程至少要经过 12 h, 但更建议在室温下进行一夜。一些蛋白质,特别是高分子质量蛋白质需要更长的时间才能在再水化过程中进入胶条内。 (3) 在完全再水化后, 将胶条移人水平电泳系统, 并将此系统的冷却块温度恒定于 20°C。 (4) 将潮湿的吸干纸铺于再水化的 IPG 胶条两端,电极放置到位,并把石蜡油覆盖在胶条之上&。 (5) 表 30.1 为典型的聚焦方案。 再水化上样是二维凝胶电泳样品导入的最简便方法,这一方法使得样品缓冲液中的蛋白质样品在 IPG 胶条吸收样品溶液时被动导入,且蛋白质可以在整个 pH 梯度中均匀分布。在一些商品化的等电聚焦仪器中,IPG 胶条的再水化和聚焦可以用相同设备仪器完成,无需人工操作。这种仪器也可以进行所谓的主动再水化,即在胶条再水化上样过程中接入低电压 (30~50V),在一些情况下, 主动再水化可以促进样品应用,因为这促使盐离子向电极移动,去除了蛋白质中的蛋白酶,并帮助大分子进入凝胶之中。然而, 等电聚焦过程中的 IPG 胶条再水化方法 (正面朝下) 有些时候也会出现较低分辨率的情况,这是由于等电聚焦胶条的多余的机械应力加在了聚焦托盘和塑料凝胶衬底之间,蛋白质数量较多时这种情况尤为常见。在这些情况下,使用聚焦仪器,并让塑料衬底直接与聚焦托盘接触 (凝胶便会正面朝上),可利于更为复杂困难的样品(图 30.3)。 (6) 可选项:主动再水化 (在一些设备配置下可以进行)。当梯度胶条正面朝下并和电极接触时,样品的再水化即可发生。再水化过程中,电极会在胶条上产生较低(通常 30~50V) 电场。在一些情况下,主动再水化能够促使高分子质量蛋白质进人梯度胶条之中。在这种配置下,等电聚焦仪器一开始聚焦于正面朝下的梯度胶条之上,而这时并不需要人工干预。 使用碱性范围的 IPG 肢进行等电聚焦再水化上样的一大优点便是省去了许多人工操作步骤。然而,它也有许多不足之处, 特别是在碱性 pH 范围中,因为在碱性环境中蛋白质会在胶条表面聚集, 并在二维凝胶中留下水平拖尾。 与酸性范围相比 JPG 胶条在碱性范围的电泳条件有所不同。在碱性 pH 环境下的等电聚焦过程,存在着还原剂 DTT 的水转运与迁移。为了将这些影响降到最低, 蛋白质样品通常在阳极一端进行杯上样, 而不是进行凝胶内再水化(图 30.3)。碱性胶的最大蛋白质载量低于酸性胶。强烈建议,每杯蛋白质上样不要超过 0.5 mg, 以防拖尾。 以下方法用于 pH3~11 和 pH7~11 的 24 cmIPG 胶条。 (1) 确保最终样品体积不超过上样杯容量 (通常为 100~120 ML, 具体参照制造商规定的容量 (2) 在溶胀盒中,IPG 胶条在含有羟乙基二磺化物 (HED) 的再水化缓冲液 (不含蛋白质样品)(DeStreak;01ssonetaL,2002) 中进行再水化。在这一阶段,可加人终浓度为 0.5% 的 IPG 胶条缓冲液,用以水平等电聚焦和垂直 SDS-PAGE。由于在再水化过程中无蛋白质存在,可将时间缩短至 6 h(为了让再水化和聚焦在同一天进行),但是通常推荐将这一时间持续在 12 h 以上。 (3) 在完全再水化后,将 IPG 胶条放置于水平电泳系统,并将此系统冷却块温度恒定于 20°C。 (4) 将潮湿的吸干纸铺于再水化的 IPG 胶条两端,电极放置到位。将上样杯小心放置在阳极 (即梯度的酸性一端) 正下方,这样一来,杯底便可直接和 IPG 胶条顶部接触并密合,但不会破坏凝胶。再将石蜡油覆盖于胶条之上。 (5) 确保石蜡油不会漏入杯中, 然后向杯中加入 1 0~2 0 的石蜡油,小心地用移液器在石蜡油下移取样品 (具体每杯最大样本容量应参照制造商规定的容量)③。 (6) 表 30.1 为典型的聚焦方案. IPG 胶的平衡等电聚焦完成后,在进行第二维电泳之前有必要先对 IPG 胶条分两个步骤进行平衡。电渗(electroendosmoticeffect) 会导致一维到二维转换的效果变差,而平衡缓冲液中含有 6mol/L 尿素和 30% 的甘油,可降低电渗的影响(Sanchezetal.,1997)。并且,平衡缓冲液中含有 2%(W/V)SDS, 使蛋白质带上负电荷以进行 SDS-PAGE。在平衡过程中及后续一维到二维的转换过程中,蛋白质的损失主要是因为蛋白质被 IPG 胶基质较强吸收以及冲洗的原因。大部分蛋白质损失都发生在平衡过程的前几分钟,而在平衡过程的第二个步骤中,蛋白质损失极少 (Sanchezetal.,1997)。 (1) 在聚焦完成之后,将石错油从 IPG 胶条上移除, 并将 IPG 胶条孵育在平衡缓冲液之中。这一步骤可以在各种容器、密封管,甚至是在平底的 IPG 胶条聚焦支持面上进行,以最大限度减少对胶条的操作。 (2) 胶条首先在 25~100 mL 的平衡缓冲液中进行平衡 (15~20 min), 缓冲液中还含有 1%~2%(m/V) 的 DDT 以减少二硫键。 (3) 将溶液倒掉, 替换成等体积含碘乙胺酰 [为 DTT 用量的 2.5 倍的平衡缓冲液,再孵育 15~20 min, 使已形成的巯基被脲甲基 (净分子质量增加 57Da) 烷基化。这样能够防止巯基在二维 SDS-PAGE 时再次氧化,从而防止蛋白质拖尾礼 (4) 这时便可将平衡后的 IPG 胶条在二维 SD&PAGE 凝胶上进行电泳,或包两层塑料膜,将其冷冻储存在一 20°C 或一 80°C 数月。对于标记 DIGE 的样本来说,以此方法储存可在至少一周内不影响 Cy 染料的性能。如有需要,可在聚焦之后,用此方法直接将 IPG 胶条冷冻,将平衡和还原/烷基化步骤留到需要二维分离时再完成。 SDS-PAGE: 第二维在平衡过程结束之后,胶条便可用于第二维 (SDS-PAGE)。当超过一块凝胶需要进行电泳时 (在比较蛋白质组学中很常见),凝胶一起进行电泳可获得最具可重复性的结果。 通常需要使用可承载 10~12 个大规格(24 cmX20 cm)SD&~PAGE 凝胶的电泳棺, 这种电泳槽在市面上可以买到。为了提高重复性、降低电泳过程中 SDS 的损耗, 建议使用冷却系统使缓冲液保持在 20°C。 自制的凝胶使用普遍、价格适宜,但需要大量人力重复制备。现成的预制凝胶在市面上也可购得, 包括用于 DIGE 的使用弱荧光支撑介质的凝胶。不同交联剂比例(丙烯酰胺:双丙烯酰胺) 的丙烯酰胺浓缩液也可使用。对于一般用途而言,12% 均一孔径凝胶 (12%T、26%C) 能够在便于制备和分辨率上达到最好的折中效果。梯度凝胶在更广的质量范围中分离效果和分辨率更好, 但却更难大规模重复制备。重复制备梯度凝胶专用的制胶器也可在市面上买到。 人工灌制的凝胶可借助多凝胶制胶器单独制作或批量制作。每份多凝胶灌制时所需要的丙烯酰胺溶液总量可根据每块独立凝胶的设置和装备提前确定。下面是终体积为 2.3L 的多凝胶制备的一个例子。若要求更小体积, 成比例地调整用量即可。 (1) 通常,用塑料板将有 1.0 mm 或 1.5 mm 厚度带有垫片的玻璃板分开,以防止凝胶聚合作用以后玻璃板之间相互粘连。如果需要,可在凝胶灌制之前在玻璃板之间放人写字的 Whatman 滤纸,以便给每块凝胶一个唯一的连续识别号码。 (2) 在真空瓶中加入 920 mL3 0%(m,V) 丙烯酰胺-双丙烯酰胺; 容液,其中丙烯酰胺!双丙烯酰胺为 37.5:1,再加入 575 mLI.5mol/LpH8.8 的 Tris-HCl 及 770 mL 水 [见步骤 (4)]。 (3) 混合物抽气处理 10 min, 以移除微小气泡, 这些气泡在某些情况下会干扰聚合作用,并在蛋白质染色后引起点拖尾。 (4) 可选项:脱气后,可将 SDS 加人到丙烯酰胺溶液中, 终浓度为 0.01%(35 mL 水中加人 2.3 g)。然而,许多研究表明,这一阶段不需加入 SDS 也可以获得品质好的溶液,最有可能是因为携带蛋白质通过凝胶的 SDS 在平衡阶段和/或电泳缓冲液作用下已经与蛋白质结合 (并且在电泳开始阶段,电场产生时,凝胶中的任何 SDS 都会在蛋白质进人凝胶前离开凝胶)。如今许多商业渠道都会提供含有或不含 SDS 的预制凝胶。若本步骤不使用 SDS,则要在步骤 (2) 中将 770 mL 加水量替换成 805 mL[见步骤 (2)]。 (5) 在灌胶之前,要采取温和方式或用一个搅拌棒混匀,在丙烯酰胺混合物中加入 7 00 mg 过硫酸铵 (APS,现用现配) 和 300TEMED(根据不同灌胶装置,丙烯酰胺混合物用量减小时,APS 和 TEMED 的用量也应成比例地减少)。 (6) 制备多凝胶所用的槽需要从底部开始浇注,至离玻璃板顶部 2 cm 的高度即可。小心地在凝胶上覆盖 I.0~I.5 mL 缓冲液饱和的异丁醇,使聚合作用开始 (7) 在聚合作用完成之后,清理制胶玻璃板上剩余的聚丙烯酰胺残留物,用水或 SDS 电泳缓冲液覆盖,用双层塑料薄膜封盖,可在 4°C 保存几周。 (8) 若先前的第一维 IPG 胶条 (已平衡和还原/烷基化) 被冷冻过,在解封和操作这些胶条前先放置几分钟使其完全解冻。冲洗并除去第二维凝胶玻璃板顶部空间中的水或 SDS 电泳缓冲液。 (9)IPG 胶条应该浸泡在 SDS 电泳缓冲液中,随后放置在 SDS-PAGE 凝胶装配的顶部孔空间内。梯度胶条背后的塑料衬片需要黏附在其中一块玻璃板的内表面 (如果可以的话通常是较长的那块玻璃板),并且随后可以简单地用薄卡片或尺子轻触放入合适位置 (确保卡片或尺子压力是作用在塑料衬片上而不是 IPG 凝胶上)。SDS 电泳缓冲液覆盖在电泳槽内后,使用同样的轻触步骤可以轻易地移除气泡,实现 IPG 胶条与二维凝胶的良好接触。装置装配完毕后,在第二维电泳运行中不可以再移动 IPG 胶条。 (10) 可选项。为确保 IPG 胶条和凝胶良好接触,可以使用琼脂糖溶液以保持 IPG 胶条处于适合位置。琼脂糖溶液需保温在 65°C 并先被加入到凝胶顶部。随后立即将平衡后的 IPG 胶条放在凝胶上面。有些人赞成这么做, 但这确实是一个笨拙的方法, 并且若在胶条放置完毕和气泡移除前琼脂糖溶液就凝固的话,将会变得难以处理。 (11) 可选项。蛋白质分子质量标准可在旁侧进行电泳 (通过小块纸片上样)。 (12) 初期将电流设置为每块凝胶 5~10mA 以使蛋白质进入凝胶,随后设置电流为每块凝胶 20mA,l5°C 过夜直至蓝色前沿到达凝胶末端。有条件的话采用恒定功率,至少在第 Ih 要将功率设置为每块胶小于 IW,随后功率每块胶不大于 15W 直到完成,这是最优设置,因为电泳时随着盐和离子离开凝胶,电压和电流强度将会发生改变。 电泳完成之后,固定凝胶并采用各种各样的可视和荧光染色技术对凝胶染色。通过荧光成伐,DIGE 胶可在第二维电泳完成后立即成像。随后可以将凝胶放在冰箱或 4°C 冷藏间里保存几个月。经二维电泳并长时间保存后的蛋白质,采用质谱分析仪来鉴定是没有任何问题的叭 展开 |