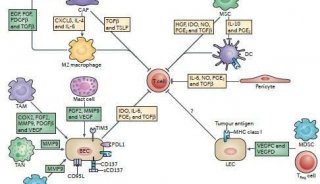
转录因子组成
真核生物在转录时往往需要多种蛋白质因子的协助。一种蛋白质是不是转录结构的一部分往往是通过体外系统看它是否是转录起始所需要的。一般这些促成转录起始所需的转录结构包括三个重要的组成部分:
亚基
RNA聚合酶的亚基,它们是转录必须的,但并不对某一启动子有特异性。
复合物
某些转录因子能与RNA聚合酶结合形成起始复合物,但不组成游离聚合酶的成分。这些因子可能是所有启动子起始转录所必需的,但亦可能仅是譬如说转录终止所必需的。但是,在这一类因子中,要严格区分开哪些是RNA聚合酶的亚基,哪些仅是辅助因子,是很困难的。
特异顺序
某些转录因子仅与其靶启动子中的特异顺序结合。如果这些顺序存在于启动子中,则这些顺序因子是一般转录结构的一部分。如果这些顺序仅存在于某些种类的启动子中,则识别这些顺序的因子也只是在这些特异启动子上起始转录必需的。
黑腹果蝇的RNA聚合酶需要至少两个转录因子方能在体外起始转录。其中一个是B因子,它与含TATA盒的部位结合。人的因子TFⅡD亦和类似的部位结合。同样,CTF(CAAT结合因子)则与腺病毒的主要晚期启动子中与CAAT盒同源的部位相结合。结合在上游区的另一个转录因子是USF(亦称MLTF),则可以识别腺病毒晚期启动子中靠近-55的顺序。转录因子Sp1则能和GC盒相结合。在SC40启动子中有多个GC盒,位于-70到-110之间。它们均能和Sp1相结合。然而含有GC盒的不同的DNA顺序与Sp1的亲和力却各不相同。可见GC盒两侧的顺序对Sp1-GC盒的结合究竟如何能影响转录。有时候需要几个转录因子才能起始转录。例如胞苷激酶的启动子需要Sp1与GC盒结合和CTF与CAAT盒结合;腺病毒晚期启动子需要TFⅡD与TATA盒结合和USF与其邻近部位相结合。以上所述的因子是一般转录都需要的,似乎并没有什么调节功能。另一些转录因子则可以调控一组特殊基因的转录。热休克基因就是一个很好的例子。真核生物的热休克基因在转录起始点的上游15bp处有一个共同顺序。HSTF因子仅在热休克细胞中有活性。它与包括热休克共同顺序在内的一段DNA相结合,所以这个因子的激活可以引起约包括20个基因的一组基因起始转录。在这里,转录因子和RNA聚合酶Ⅱ之间关系很类似细菌的σ因子与核心酶之间的关系。
转录因子是一种具有特殊结构、行使调控基因表达功能的蛋白质分子,也称为反式作用因子。植物中的转录因子分为二种,一种是非特异性转录因子,它们非选择性地调控基因的转录表达,如大麦(Hordeum vulgare)中的HvCBF2(C-repeat/DRE binding factor 2)(Xue et al., 2003)。还有一种称为特异型转录因子,它们能够选择性调控某种或某些基因的转录表达。典型的转录因子含有DNA结合区(DNA-binding domain)、转录调控区(activation domain)、寡聚化位点(oligomerization site)以及核定位信号(nuclear localization signal)等功能区域。这些功能区域决定转录因子的功能和特性(Liu et al., 1999)。DNA结合区带共性的结构主要有:1)HTH 和 HLH 结构:由两段α-螺旋夹一段β-折叠构成,α-螺旋与β-折叠之间通过β-转角或成环连接,即螺旋-转角-螺旋结构和螺旋-环-螺旋结构。2)锌指结构: 多见于 TFIII A 和类固醇激素受体中,由一段富含半胱氨酸的多肽链构成。每四个半光氨酸残基或组氨酸残基螯合一分子 Zn2+ ,其余约 12-13 个残基则呈指样突出,刚好能嵌入 DNA 双螺旋的大沟中而与之相结合。3)亮氨酸拉链结构:多见于真核生物 DNA 结合蛋白的 C 端,与癌基因表达调控有关。由两段α - 螺旋平行排列构成,其α - 螺旋中存在每隔 7 个残基规律性排列的亮氨酸残基,亮氨酸侧链交替排列而呈拉链状,两条肽链呈钳状与 DNA 相结合。