植物膜受体如何在共生和免疫信号之间一碗水端平
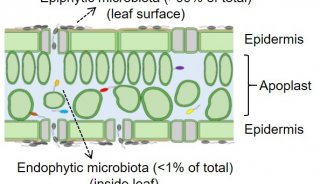
在整个生命周期中,植物必须对各种微生物作出适当的反应。在与无害的共生体一起生活并抵御致病和寻求营养的病原体的同时,植物还与提供稀缺矿物质营养的微生物进行亲密的内共生(Science | 重磅研究揭示植物如何区分有益和有害微生物!Cell | 瑞士洛桑大学研究揭示植物根部免疫系统如何区别对待病原微生物和有益微生物!Cell Host & Microbe | 植物根部采用细胞层特异性程序响应病原微生物和有益微生物!Nature Plants | 德国马普植物育种所研究揭示植物先天免疫协调微生物-宿主的平衡!)。特别是,与丛枝菌根真菌(AMF)的互利共生非常普遍,并被认为有助于植物在大约4.5亿年前对陆地的定殖。大约在4.5亿年前,它对植物的定殖起到了重要作用(Science | 植物与真菌的海誓山盟!研究揭示脂质交换驱动植物陆地化过程中的共生进化!Science | 重磅!植物和微生物的新途径:与共生微生物和病原微生物的相互作用驱动植物进化!)。直到今天,它对植物营养仍然很重要。植物决定与适当的微生物接触的分子机制是一个热点研究的领域,收集到的知识可以使作物的发展更多地受益于其微生物组,并减少其损害。
近日,国际权威学术期刊PNAS发表了英国剑桥大学Chai Hao Chiu和Uta Paszkowski的最新相关研究成果,题为How membrane receptors tread the fine balance between symbiosis and immunity signaling的评论文章。


植物识别微生物的最早步骤之一是感知植物中没有的保守分子,也被称为微生物相关分子模式(MAMPs)。20多年来,植物受体已经被确认,可以识别它们的同源MAMPs并转发细胞信号(Cell Host & Microbe | Jeff Dangl团队揭示共生菌群通过MAMP多样性调节植物免疫!Cell Host & Microbe | 背靠背!Jeff Dangl团队揭示拮抗多效性调节植物免疫的机制!Cell Host & Microbe | 细菌鞭毛蛋白与植物免疫系统之间的 “猫鼠游戏”!Plant Cell | 牛津大学阐述植物病原菌和共生菌避免细胞表面受体识别的九种胞外策略!)。这些受体识别保守的MAMPs,从肽(如鞭毛蛋白表位)到碳水化合物(如微生物细胞壁成分),是植物先天免疫监视系统的典型。对碳水化合物MAMPs(如几丁质或肽聚糖)的感知涉及到质膜上的溶素基序受体类蛋白(RLPs)或受体类激酶(RLKs)。最早在噬菌体中发现的溶素基序是一个碳水化合物结合域,对N-乙酰葡糖胺(GlcNAc)的聚合物有亲和力,后者形成几丁质,是肽聚糖的一个组成部分,而根瘤菌的结节因子也被称为脂壳寡糖(LCOs)。长链壳寡糖(COs),如壳寡糖(CO8)和肽聚糖,通常被认为是与病原体相关的,而短链壳四糖(CO4)和LCOs是与共生体相关的MAMPs,当植物与病原体和共生体接触时,正是这些MAMPs被感知到。
来自不同植物物种的证据表明,LysM-RLKs/RLPs在免疫和共生背景下的碳水化合物感知中发挥了作用。LysM-RLKs结瘤因子受体1(NFR1)和NFR5在Lotus japonicus的正向突变体筛选中被确定为感知LCO(也被称为结瘤因子)以启动与根瘤菌固氮共生的信号所必需。在水稻中,亲和标记确定几丁质诱导子结合蛋白(CEBiP)是主要的CO结合受体。CEBiP和它的核心受体Chitin Elicitor Receptor Kinase 1(CERK1)是几丁质触发的免疫所必需的。有趣的观察是,水稻CERK1突变体显示出丛枝菌根真菌共生的减少,这表明对病原体发出的信号和促进共生的因子(也被称为myc因子,类似于根瘤菌结瘤因子)的感知涉及一个共享的核心受体。事实上,我们现在知道CERK1是碳水化合物MAMP感知中的一个主要核心受体,并且在迄今为止测试的大多数(但不是全部)植物物种中参与免疫和菌根共生信号。
因此,关于植物如何区分共生体和病原体的一个有吸引力的模型是,不同的CERK1-核心受体复合物的形成导致不同的信号输出。在PNAS上,Zhang等人(PNAS | 中科院王二涛团队揭示受体竞争可以区分水稻的共生和免疫信号!)现在提供证据表明,根据招募CERK1的受体,水稻植株区分共生与免疫信号,如图所示。有丝分裂原激活蛋白激酶(MAPKs)的磷酸化和活性氧(ROS)的产生都代表了信号转导级联的激活,并经常被用作免疫信号的输出,而核钙振荡则作为与共生有关的反应的代理。Zhang等人使用纯化的COs表明,CO8在激活MAPK和ROS的同时也激活了与共生有关的核钙振荡,而CO4只引起了核钙振荡。这些数据表明,植物对AMF的早期感知很可能涉及CO8,也可能涉及CO4,这证实了其他人的报告。
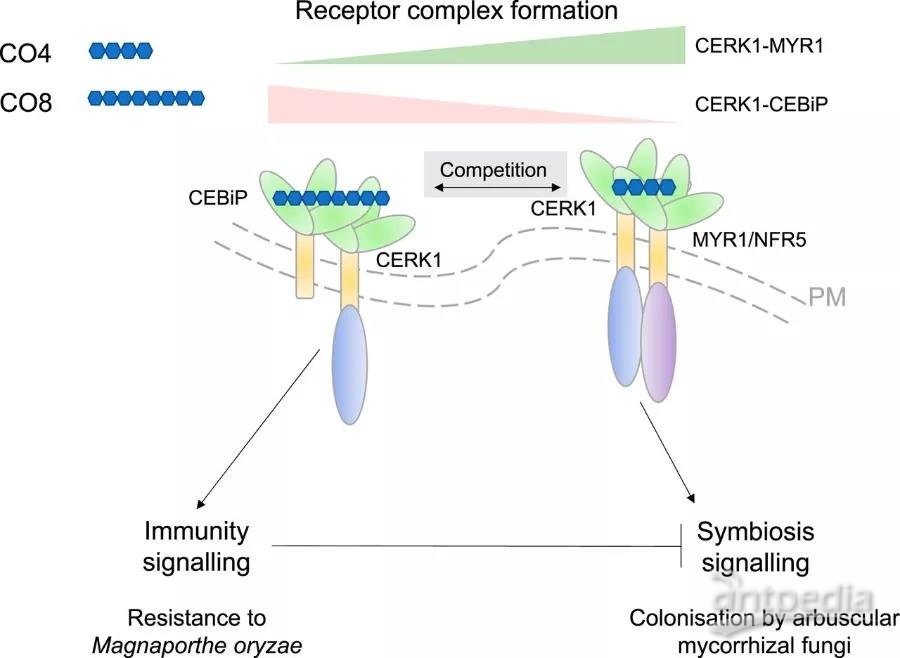
观察到用CO4预处理可以抑制CO8诱导的ROS,并确认这种抑制需要CO4结合受体--Myc Factor Receptor 1,以前也被称为Nod Factor Receptor 5(MYR1/NFR5),暗示了质膜上复杂受体动态的可能性。在拟南芥原生质体中进行的一系列生化实验表明,CO4减弱了CERK1-CEBiP的相互作用,而MYR1的存在又进一步增强了这种作用。与生化证据一致,myr1突变体的生理表型包括更高水平的CO8诱导的ROS,防御基因的表达增加,以及稻瘟病真菌Magnaporthe oryzae的定殖减少。这些试验共同支持这样的观点:在没有MYR1的情况下,CERK1-CEBiPe的免疫反应更强。
在过量表达MYR1-GFP的植物中,水稻对稻瘟病真菌的敏感性增强,这进一步证明了相互作用和免疫力之间的微妙平衡。正如本研究和早期工作所证明的,在感知AMF时,免疫和共生信号反应都被激活。因此,免疫受体CEBiP的损失预计会增加由CERK1-MYR1复合体所施加的共生信号反应。因此,Zhang等人注意到cebip突变体的菌根共生微妙增加,这发生在共生的早期阶段(接种后2周),此后便消失了。由于可以预料到在整个共生体建立过程中不断需要控制免疫信号,因此在没有免疫激活的CERK1-CEBiP复合体的情况下,共生体的适度和短暂的促进是一个有待进一步研究的谜团。
综上所述,Zhang等人的工作为一个模型提供了证据,其中CERK1-CEBiP复合体支持免疫反应,而CERK1-MYR1/NFR5复合体在有益的AMF存在时激活共生信号。因此,受体复合体之间的竞争导致了共生和免疫的对抗性信号输出--如图所总结。
然而,还有许多悬而未决的问题。例如,不同的几丁质信号-CO4、CO8和LCOs在被植物感知时是如何整合的?我们现在了解到,广泛的真菌(包括担子菌和子囊菌)产生LCOs,这种化合物通常被认为是一种共生信号,因为它事先被确认为根瘤菌的结瘤因子。另一方面,几丁质是真菌细胞壁不可缺少的成分,也是丰富的植物几丁质酶的底物,其活性可产生包括CO4和CO8在内的COs。这样看来,根际不可避免地富含CO8、CO4和LCOs。CO4和LCOs都能够减弱CO8引起的早期免疫相关的ROS和MAPK激活。所有这些几丁质信号都能激活核钙振荡(共生信号的一个标志),但在这项工作中,CO8最终抑制了与AMF的共生。Zhang等人将他们的工作重点放在CO4上,提到LCOs不被水稻感知,尽管值得注意的是,作者以前证明了在水稻中较高浓度的LCOs下发生核钙振荡,这与在其他植物物种中的观察结果一致。
另一个重要的未回答的问题与已知的受体配体杂乱性有关,不严格地与CO4或CO8结合,以及如何将其纳入模型。这项工作和以前的工作表明,CEBiP与CO4和CO8结合的亲和力相似,而MYR1与CO4结合,与CO8的亲和力较低。那么,与CO4和CO8结合的受体复合体的相对数量及其组成是什么?显然,需要做更多的工作来了解植物究竟如何利用CO8、CO4和LCO的感知来区分有益的和致病的微生物。本文中介绍的受体竞争提供了一种机制,标志着我们开始更全面地了解植物界如何感知陆地第二丰富的聚合物,以及植物如何与适当的微生物形成共生关系。