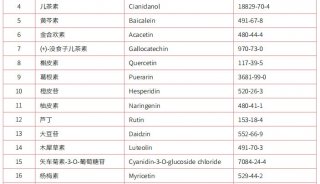
上海百趣生物医学科技有限公司

 分享至
分享至



百趣代谢组学资讯:机制探索不发愁,浅看真菌防治靶点代谢组学研究思路
文章标题:Energy metabolism as the target of3-phenyllactic acid against Rhizopus oryzae
发表期刊:InternationalJournal of Food Microbiology
影响因子:5.277
合作客户:兰州理工大学
百趣生物提供服务:TMT+中心碳
分组:2mg/mLPLA治疗组、4mg/mLPLA治疗组、8mg/mLPLA治疗组和对照组(n=3)
研究背景
3-苯乳酸(PLA)具有广泛的抗真菌活性,但对PLA在真菌细胞上的作用靶点及其作用机制的研究较少。本研究探讨了3-苯乳酸对引起腐烂百合鳞茎的米根霉菌(R.oryzae)的抑制作用。PLA对米根霉菌的最低抑菌浓度为8mg/mL。通过扫描电镜和透射电镜观察米根霉菌的超微结构,结果表明PLA对米根霉菌的细胞膜没有损伤,但对线粒体和其他细胞器有破坏作用。TMT蛋白组分析结果显示PLA显著下调己糖激酶(HK)、磷酸果糖激酶(PFK)、a-酮戊二酸脱氢酶(a-KGDH)、腺苷酸激酶(ADK1)、细胞色素C氧化酶(Cytochrome C oxidase)和NADH脱氢酶(NADH dehydrogenase)的表达(P<0.05)。在糖酵解、三羧酸循环或氧化磷酸化代谢过程中,线粒体ADP/ATP载体蛋白(AAC)和亚基IV(CCIO IV)表达上调(P<0.05)。在此基础上,通过酶活性检测发现上述通路HK和a-KGDH活性下调,并通过实时荧光定量PCR进一步证实ADK1、AAC、CCIO IV和NADH脱氢酶基因表达下调。中心碳代谢组学研究表明,三羧酸循环代谢产物中柠檬酸、顺-乌头酸、异柠檬酸、-酮戊二酸、琥珀酸、富马酸和苹果酸均显著下调(P<0.05),氧化磷酸化产生的ATP也显著减少(P=0.02)。导致能源生产不足。因此,ROS水平升高了141%,细胞色素C被释放,细胞逐渐凋亡。结果表明,能量代谢是PLA对米根霉的作用靶点。这一研究首次表明PLA对米根霉的能量代谢可能是其作用靶点,为进一步研究其抑菌机理提供了理论依据。
百趣代谢组学:机制探索不发愁,浅看真菌防治靶点代谢组学研究思路
研究结果
3-苯乳酸对米根霉生长的抑制作用
PLA处理组(2和4mg/mL)菌体的菌丝干重均低于对照组。当PLA浓度达到8 mg/mL时,随着培养时间的延长,米根霉测得的干重基本没有变化,说明在该浓度下米根霉的生长受到了严重的抑制。实验结果表明,3-苯乳酸对米根霉的生长有较好的抑制作用,且随着3-苯乳酸浓度的增加,对米根霉的抑制作用越明显。

图1. 米根霉的生长曲线(灰色线为对照组,红色线为2mg/mLPLA治疗组
3-苯乳酸对菌丝形态和超微结构的影响
用扫描电镜(SEM)观察了稻瘟菌的菌丝形态。对照样品形态正常,菌丝饱满,表面光滑(图2.A1和A3)。PLA处理米根霉24h后,我们发现菌丝密集杂乱(图2.A2和A4),菌丝管肿胀起皱;但表面无明显损伤或缺失(图2.A4)。在菌丝表面观察到一些分泌物(圆圈),推测为胞外多糖(图2.A4)。因此,推断PLA处理对米根霉细胞壁没有造成严重的破坏。
电镜观察显示,对照组米根霉的超微结构完整,细胞壁正常光滑,细胞膜结构正常,胞浆壁无分离,线粒体形成相对规则(图2.B1-B4)。经3-苯乳酸处理后,整个细胞结构萎缩,细胞外壁变薄,厚度不均匀。内侧细胞膜呈齿状凹陷,局部浆壁分离。细胞核消失,基质和内嵴不可见。细胞膜呈齿状向内,浆壁部分分离(图2.B4中红色箭头)。线粒体固缩,液泡塌陷。
综上所述,PLA对米根霉线粒体及其他细胞器的破坏程度较高,这可能是PLA对米根霉抑制作用的靶点。

图2.对照和PLA处理米根霉的SEM和TEM检查:(A)SEM检查,(B)TEM检查
3-苯乳酸的蛋白表达对米根霉菌有抑制作用
利用TMT蛋白质组学方法,在PLA处理组和对照组之间发现了485个差异表达的蛋白。其中,PLA处理组有205个蛋白表达上调,280个蛋白表达下调。对差异表达蛋白进行COG分析(图3)得知PLA可通过控制能量代谢影响米根霉菌的生长。且经PLA处理后的米根霉菌中HK、FK、SDH和a-KGDH的活性受到抑制。相比之下,PLA处理后的米根霉菌中PFK活性略有增加,碳流可能被转移到其他丙酮酸分支点。这些结果与差异蛋白表达是一致的。

图3. 差异表达蛋白的COG分析

图4. 与糖酵解和TCA循环相关的关键酶活性,星号表示样本间差异显

表1. 4mg/mLPLA处理组与对照组在糖酵解、三羧酸循环或氧化磷酸化
米根霉菌中心碳代谢产物的差异及分析
通过中心碳检测,我们在对照组和PLA处理组之间确定了51个中心碳代谢产物,其中22个代谢产物显著上调,29个代谢产物显著下调 (图6)。利用KEGG数据库对差异代谢产物的代谢途径进行分析,差异表达的代谢产物在TCA循环、乙醛酸和二羧酸代谢、糖酵解或糖异生、丙酮酸代谢、光合生物碳固定和磷酸戊糖途径中显著富集。在本研究中, PLA处理显著降低了米根霉菌中ADP和ATP的水平(表2),说明PLA处理影响了米根霉菌的能量代谢。机体线粒体中的OXPHOS负责90%以上的细胞ATP的产生,结合蛋白质组学分析,这种效应可能是由于OXPHOS中一种或多种酶活性明显缺乏,导致ATP合成减少。

表2. 4mg/mLPLA处理组与对照组在糖酵解、三羧酸循环或氧化磷酸化

图6. 中心碳代谢产物在对照组和PLA处理组之间的差异
3-苯乳酸对细胞色素C释放和ROS水平的影响
经过PLA处理的米根霉菌细胞质中的细胞色素C含量更高(图7A),这表明位于线粒体内外膜的细胞色素C缺失。在之前的一项研究中,反式肉桂醛处理导致细胞色素C渗漏并触发线粒体介导的细胞凋亡。推断PLA可能导致线粒体呼吸链功能障碍,进而导致ATP缺乏,导致细胞死亡。
当PLA处理米根霉菌,ROS水平增加,在4.0mg/LPLA时,ROS水平增加了141%(图7B)。结果表明,PLA处理诱导水稻植株ROS积累,可能引起线粒体功能障碍和细胞氧化损伤。
在本研究中,我们发现PLA影响糖酵解、TCA循环和氧化磷酸化,导致能量生产不足和细胞凋亡。虽然目前还没有关于对米根霉菌的抑制作用的研究,但与乳酸相比,PLA对其他真菌如黑曲霉具有显著的抑制作用,与乙酸相比,PLA没有明显的刺激性气味。此外,使用乳酸作为催化剂生产PLA可能是一种可行且环保的方法。因此,PLA可以作为一种很有前景的天然机制来控制食品中的污染物。

图7. 对照组和PLA处理组之间的ROS水平和Cyt C释放,星号表示样本
结论
PLA对真菌细胞靶点的作用及其抗真菌机制研究较少。本研究通过生理生化、电镜、TMT蛋白质组学和中心碳代谢组学等方法,探讨了米根霉菌对百合腐烂球茎的抑制作用。首先,我们证明了PLA抑制了米根霉菌的生长,破坏了米根霉菌的形态和超微结构。其次,我们发现PLA处理降低了与糖酵解和TCA循环相关的关键酶的活性,导致丙酮酸和TCA循环产物的减少和能量的损失。最后,PLA处理通过影响电子传递链中的关键蛋白来影响氧化磷酸化,导致能量生产不足,且使米根霉菌的ROS水平升高,细胞色素C释放,导致细胞凋亡逐渐发生。综上所述,PLA通过能量代谢抑制米根霉菌的生长(图8)。这是首次对PLA抑制真菌的机制进行全面研究。因此,PLA可作为一种有效的天然抗真菌药物。

图8. PLA抑制稻瘟病菌的可能机制是基于能量代谢,紫色代表减少,
文/阿趣代谢组学
-
厂商文章

-
厂商文章
-
厂商文章
-
厂商文章
-
厂商文章

-
厂商文章
-
厂商文章
-
厂商文章

-
厂商文章

-
厂商文章

-
厂商文章
-
厂商文章

-
厂商文章
-
厂商文章
-
厂商文章
-
厂商文章
-
厂商文章

-
厂商文章