Nature综述丨冷冻电镜在药物发现中的应用前景
6药物筛选中X-ray晶体学与Cryo-EM的比较
与Cryo-EM相比,X-ray晶体学在药物筛选方面的关键优势在于,能够快速提供高分辨率结构数据。一旦建立了合适的结晶系统,通过X-ray晶体学快速连续获得后续结构或筛选化合物所需的时间非常短:1小时内可以通过结晶自动工作站设置约2,000个结晶条件,2-3个小时即可评估所有图像以确定合适的结晶条件(可以通过图像识别软件来降低)。在同步加速器上采集一套完整的晶体数据不到2分钟,随后的数据整合、结构解析和蛋白质-配体复合物的精修已经很大程度上已经实现自动化,并且通常可以在1小时内完成。然而,筛选以确定EM的合适样品和冷冻条件时,1次分析仅能评估12个样品,而为了充分表征它们又需要3-4个小时,与X-ray相比慢了好几个数量级。
重要的是,这意味着X-ray晶体学的通量对于药物配体筛选足够快。目前,用不同化合物浸泡的约500个晶体的数据可以在24小时内收集(例如,在英国Diamond synchrotron的XChem线站;上海SSRF的BL17U和BL19U1也有此潜力),并且可以在在1周内在线处理和精修好(视结构解析者的数量和熟练程度)。 为了实现相同的目标,EM则需要大约半年的时间收集数据(假设每套数据8小时),并且至少需要一年的计算和模型构建。即使有这些乐观的时间尺度假设,使用Cryo-EM的配体筛选过程也比使用X-ray晶体学的要慢2-3个数量级。
正如前文指出的那样,cryo-EM的一个关键优势是:它可以很容易地用于确定其天然状态下的大分子和/或动态大分子(包括膜蛋白)的结构,包括翻译后修饰。Cryo-EM对蛋白质的需求通常比X-ray晶体学更少,这对于要求苛刻的蛋白质可能是特别有利的。事实上,对于在药物研发中使用X-ray晶体学,获得合适的晶体仍然是一个关键挑战,有延迟项目的风险,特别是膜蛋白,重糖基化蛋白和大型或柔性蛋白或蛋白复合物,能否持续获得稳定、可靠、质量上乘的晶体存在很多不确定因素。为了应对这些挑战,人们已经开发了许多方法,例如将大的药物靶标减少到单个结构域,以期促进蛋白质生产,纯化和结晶。所以,在结构化指导的药物发现项目开始时,对时间和资源的投入往往是强制性的,以确保在筛选数百种化合物时具有足够的通量。
Cryo-EM的另一个重要优点是相位信息可以在重建结构和实验中直接获得,而相位信息在X-ray晶体学中丢失,必须通过实验进行相位重构,这取决于实验数据的准确性采集。因此,尤其是对于4-7.5 Å范围内的分辨率水平,通过Cryo-EM确定的结构,通常能够以无法通过X-ray晶体学获得的清晰度显现单个结构域和二级结构单元。但是,分辨率超过3 Å时情况不同。只有一小部分已发表的Cryo-EM结构有超过3 Å的分辨率,并且在许多情况下,从Cryo-EM获得的3D图谱无法在质量上与通过X-ray晶体学获得的图谱在类似的分辨率下竞争。不过,在这一方面应该指出的是,X-ray晶体学和Cryo-EM的分辨率基于的方法差异很大,因此很多时候不能直接比较(见下一部分)。
7X-ray晶体学和Cryo-EM的分辨率比较
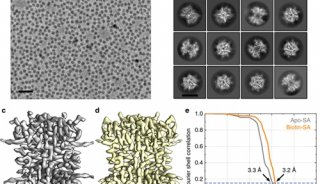
在X-ray晶体学中,基于晶格的光子最大衍射、统计学、电子密度图谱与原子模型比较等标准直接度量分辨率【27】。在Cryo-EM中,傅里叶壳层相关函数(FSC)通常被用作主要的分辨率确定工具,用两个独立确定的三维图谱的一致性来衡量两个独立确定的三维图谱。分辨率由相关性下降到特定阈值(通常为0.143)以下的空间频率确定。FSC代表数据处理的自我一致性测量;因此,在Cryo-EM密度图中观察到的结构特征有时可能会偏离他们应该看到的分辨率期望。下图中显示了这一点,其中包含一系列Cryo-EM图,显示了在不同模拟和实际计算分辨率下可视化的芳香族和非芳香族残基(图5)。
图5. 芳香族和非芳香族残基的EM密度图
FSC是一种全局措施,因此仅提供一个平均分辨率值,考虑到大分子的结构柔性和典型的Cryo-EM辐射图中的非各向同性分辨率分布,这其实是误导性的。在最佳定义的密度图区域中,分辨率可能被低估;而所有Cryo-EM密度图的结构特征比FSC所指示的分辨率低得多。同样值得注意的是,FSC没有提供关于计算3D图谱给定质量的信息。
目前,有许多提交数据库的Cryo-EM结构报道有约3.5 Å的分辨率,与3.5 Å分辨率的X-ray晶体结构相比,它们的结构特征更为明确(在其结构明确的区域)。这部分是因为如上所述,FSC的全局分辨率低估了明确界定的特征分辨率。然而,这也是因为在分辨率>3.5 Å时,X-ray晶体学的结构解析在技术上是很困难的。其原因在于X-ray晶体学的观测参数比较差。因此,从采集的幅度数据进行相位重构效率低下,这反映在这些分辨率下的差的数据质量上。当晶体学数据处理变得越来越自动化和可靠时,这种情况在较高分辨率下会反转。相比之下,与3.5-4 Å分辨率范围内的结构相比,更高分辨率的结构测定(<3Å分辨率)在Cryo-EM中困难得多。Cryo-EM结构测定中有几个参数可能会导致这种影响。例如,区分可能属于给定大分子复合物不同构象的粒子图像是非常困难的--随着一个样品中存在不同状态的数量,这个问题变得越来越突出。诸如光束损伤和电子光学像差等其他效应也越来越限制高分辨率结构测定(<2.5Å)。当想确定<2 Å分辨率下的可靠结构时,轴向彗形像差将成为Cryo-EM中主要的分辨率限制像差。
8Cryo-EM技术的未来展望
8.1目前Cryo-EM技术的瓶颈
处理较小的蛋白。Cryo-EM一直被认为是只能适用于分子量大于500 kDa的生物大分子,然而,很多的药物靶点都是比这小的多的蛋白或者蛋白复合物。近年来技术的发展,理论上可以使Cryo-EM的样品小于65 kDa,实际上也确实在这方面有很多很大的突破。因此,越来越多的小于200 kDa的高分辨结构被解析出来。
样品制备的优化和数据分析。尽管Cryo-EM对样品的均一性没有X-ray那么高,但是样品的均一性还是保持样品完整度的重要保证。不均一的样品对后期数据处理非常艰难,而且也有可能使得数据的分辨率和结构信息的完整度下降。同样,EM Grid(格栅)的准备过程、数据采集和数据等过程的优化也可以提高Cryo-EM的分辨率。随着科学家的不懈努力,相信Cryo-EM技术会越来越强大,应用会越来越广泛。
8.2Cryo-EM技术的未来
在短短的几年内,令人振奋的Cryo-EM技术的进步推动了一个时代。今天,结构生物学家正在使用cryo-EM研究大型蛋白质复合物,大型细胞机器和病毒的结构和功能。技术的改进可能很快会产生更多小的膜蛋白结构。这些进步推动了人们的希望,未来的发展可能很快就会在“可以常规实现2 Å分辨率的黄金时代”即将来临时出现【4】,并且甚至提高了期待——Cryo-EM快速而有效的预期将与许多应用竞争,甚至取代晶体学【2, 3】。
事实上,对于制药行业的一些应用,Cryo-EM已经具有相当大的吸引力,因为它可能并不需要总是达到4 Å以下的分辨率;低分辨率结构对于更好地理解目标蛋白,以及位于蛋白质复合物的结构域或结合伴侣之间结合口袋的识别;药物设计时也可能不总是需要准确确定化合物的结合模式。此外,尽管Cryo-EM结构的分辨率通常受蛋白质动态变化导致的构象多样性限制,但由于重构各种中间状态有助于理解SBDD,所以该限制可转化为SBDD的优势,并解释一个复合体如何与其底物结合的机制。
为了利用这些机会,很多公司已经将更多的注意力集中在Cryo-EM上,使用各种方法来收集和处理EM数据,包括与学术实验室合作(通过合作或收费服务)。虽然即将推出的类似于同步加速器的设备承诺满足对Cryo-EM日益增长的需求,但存在诸多限制。在晶体学方面,晶体数据收集,结构解析与优化都非常迅速,使其成为需要多种结构小分子项目的极佳资源。在Cryo-EM中,每个数据采集跨越几个小时或几天的时间,使得cryo-EM的通量比晶体学小得多。但可以肯定的是,工业中使用Cryo-EM的速度正在迅速增加。因此,对于常规的药物研发来说,仍需要开发一些工具,才能使Cryo-EM成为药物猎手的首选方法----这需要投入强大的资金和耐心。
最后,在药物研发中使用Cryo-EM不仅取决于Cryo-EM技术的的发展,还可能取决于晶体学的进一步发展。在今天,可以看到完善的、自动化的、快速的、易于量化的晶体结构筛选体系,因为这个领域已经发展了几十年的时间。很难想象在接下来的几年或几十年中,Cryo-EM会发展成什么形式和速度,可以说,取代晶体学用于常规药物发现也不是没有可能。不过,可能更好的、更有可能的结果是:虽然选择二者的界限已经开始转向有利于Cryo-EM的方向,但似乎更有可能这两种技术仍将作为药物发现的方法、互相补充共同进步很长一段时间。
本文主要内容整理自参考文献【1, 5, 6】,可能存在个别专业词汇翻译不够准确的情况,敬请谅解!有兴趣的读者请查看原文。
参考文献
1. Renaud, J.P., et al., Cryo-EM in drug discovery: achievements, limitations and prospects.Nat Rev Drug Discov, 2018.
2. Luo, F., et al., Atomic structures of FUS LC domain segments reveal bases for reversible amyloid fibril formation.Nat Struct Mol Biol, 2018. 25(4): p. 341-346.
3. Rubinstein, J.L., Cryo-EM Captures the Dynamics of Ion Channel Opening.Cell, 2017. 168(3): p. 341-343.
4. Frank, J., Advances in the field of single-particle cryo-electron microscopy over the last decade.Nat Protoc, 2017. 12(2): p. 209-212.
5. Smith, J.S., R.J. Lefkowitz, and S. Rajagopal, Biased signalling: from simple switches to allosteric microprocessors.Nat Rev Drug Discov, 2018. 17(4): p. 243-260.
6. Hauser, A.S., et al., Trends in GPCR drug discovery: new agents, targets and indications.Nat Rev Drug Discov, 2017. 16(12): p. 829-842.
7. Liang, Y.L., et al., Phase-plate cryo-EM structure of a class B GPCR-G-protein complex.Nature, 2017. 546(7656): p. 118-123.
8. Zhang, Y., et al., Cryo-EM structure of the activated GLP-1 receptor in complex with a G protein.Nature, 2017. 546(7657): p. 248-253.
9. Liang, Y.L., et al., Phase-plate cryo-EM structure of a biased agonist-bound human GLP-1 receptor-Gs complex.Nature, 2018. 555(7694): p. 121-125.
10. Bai, X.C., et al., An atomic structure of human gamma-secretase.Nature, 2015. 525(7568): p. 212-217.
11. Cao, E., et al., TRPV1 structures in distinct conformations reveal activation mechanisms.Nature, 2013. 504(7478): p. 113-8.
12. Liao, M., et al., Structure of the TRPV1 ion channel determined by electron cryo-microscopy.Nature, 2013. 504(7478): p. 107-12.
13. Gao, Y., et al., TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action.Nature, 2016. 534(7607): p. 347-51.
14. Paulsen, C.E., et al., Structure of the TRPA1 ion channel suggests regulatory mechanisms.Nature, 2015. 525(7570): p. 552.
15. Wu, J., et al., Structure of the voltage-gated calcium channel Ca(v)1.1 at 3.6 A resolution.Nature, 2016. 537(7619): p. 191-196.
16. Shen, H., et al., Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution.Science, 2017. 355(6328).
17. Hirschi, M., et al., Cryo-electron microscopy structure of the lysosomal calcium-permeable channel TRPML3.Nature, 2017. 550(7676): p. 411-414.
18. Guo, J., et al., Structures of the calcium-activated, non-selective cation channel TRPM4.Nature, 2017. 552(7684): p. 205-209.
19. Yan, Z., et al., Structure of the rabbit ryanodine receptor RyR1 at near-atomic resolution.Nature, 2015. 517(7532): p. 50-55.
20. des Georges, A., et al., Structural Basis for Gating and Activation of RyR1.Cell, 2016. 167(1): p. 145-157 e17.
21. Johnson, Z.L. and J. Chen, Structural Basis of Substrate Recognition by the Multidrug Resistance Protein MRP1.Cell, 2017. 168(6): p. 1075-1085 e9.
22. Long, F., et al., Cryo-EM structures elucidate neutralizing mechanisms of anti-chikungunya human monoclonal antibodies with therapeutic activity.Proc Natl Acad Sci U S A, 2015. 112(45): p. 13898-903.
23. Ciferri, C., et al., The trimeric serine protease HtrA1 forms a cage-like inhibition complex with an anti-HtrA1 antibody.Biochem J, 2015. 472(2): p. 169-81.
24. Wong, W., et al., Cryo-EM structure of the Plasmodium falciparum 80S ribosome bound to the anti-protozoan drug emetine.Elife, 2014. 3.
25. Wong, W., et al., Mefloquine targets the Plasmodium falciparum 80S ribosome to inhibit protein synthesis.Nat Microbiol, 2017. 2: p. 17031.
26. Fitzpatrick, A.W.P., et al., Cryo-EM structures of tau filaments from Alzheimer's disease.Nature, 2017. 547(7662): p. 185-190.
27. Karplus, P.A. and K. Diederichs, Linking crystallographic model and data quality.Science, 2012. 336(6084): p. 1030-3.