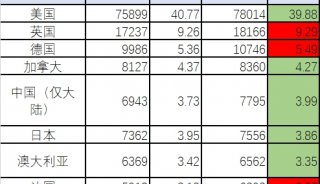
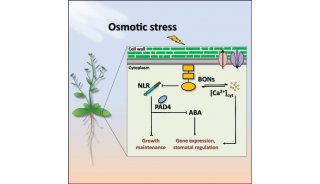
朱健康院士PLOS最新研究成果
植物激素脱落酸(ABA)调节着植物的生长、发育和对生物/非生物胁迫的响应。核心的ABA信号通路是由三个主要部分组成:ABA受体(PYR1/PYLs)、2C型蛋白磷酸酶(PP2C)和SNF1相关蛋白激酶2(SnRK2)。然而,ABA信号的复杂性,仍然是亟待解决的问题。
最近,国际遗传学期刊《PLOS Genetics》在线发表了中科院上海生命科学研究院朱健康院士带领的一项研究,题为“Type One Protein Phosphatase 1 and Its Regulatory Protein Inhibitor 2 Negatively Regulate ABA Signaling”。这项研究将TOPP1和AtI-2作物ABA信号转导的负调节因子。
朱健康是国际著名植物生物学家、植物抗逆分子学领军科学家,美国科学院院士、美国普渡大学生物化学系和园艺及园林系杰出教授,首批“千人计划”入选者,中科院上海植物逆境生物学研究中心主任。朱健康1988年赴美留学,2000年受聘美国亚利桑那大学植物科学系正教授,曾任加州大学河滨分校整合基因组学研究所所长。在植物抗旱、耐盐与耐低温方面做出了杰出成就,是世界植物科学领域发表论文引用率最高的科学家之一。近几个月发表多项重要学术成果:朱健康教授发表PNAS转基因研究新成果;朱健康院士:开发新型多路复用CRISPR/Cas9平台;朱健康院士Cell Research发布表观遗传新发现。
ABA控制着植物的多个发育过程,如种子休眠、萌发,根/茎的生长,开花和衰老。当植物遇到压力的情况,ABA可以迅速诱导的基因表达的重新编程,并触发多个生理反应,如气孔关闭,以减少植物的水分损失。鉴于ABA在调节植物生长各个方面和应激反应中的重要性,了解ABA响应不利环境条件的分子机制,是至关重要的。
拟南芥ABA abi1-1和abi2-1突变体,是在ABA不敏感表型的遗传筛选中分离出来的。这两种突变体在种子萌发、幼苗生长和气孔关闭过程中,均表现出显性负效应。有研究发现,ABI1和ABI2编码同源2C型丝氨酸/苏氨酸(Ser/Thr)磷酸酶(PP2Cs),表明磷酸酶介导的去磷酸化对于调节ABA信号起着关键的作用。据表明,这些clade A PP2Cs负向调控ABA信号转导通路。
SNF1相关蛋白激酶(SnRK2s)也调节ABA信号转导。拟南芥突变体ost1-1/snrk 2.6(Open Stomata 1,又名SnRK2.6)表现出:ABA诱导的气孔关闭受损,光诱导的气孔开放也出现缺陷。在拟南芥中已确定了10个SnRK2(SnRK2.1–10);然而在原生质体的瞬时表达检测中,只有4个(SnRK2.2 / 2.3 / 2.6 / 2.8)可以通过ABA激活,从而表明,SnRK2成员可能在依赖或不依赖ABA的信号转导途径中发挥作用。ABA诱导的基因表达和其他ABA反应,在snrk2.2/3/6三突变体植株中几乎是完全阻断的,从而表明这三个SnRK2s是ABA信号转导关键的正向调节因子。结果表明,分支A PP2Cs与SnRK2s活化环上的SnRK2s和去磷酸化关键Ser/Thr残基结合。
近年来,ABA受体被确定为PYRABACTIN RESISTANCE 1 (PYR1)/PYR1-Like (PYL)/REGULATORY COMPONENT OF ABA RECEPTOR (RCAR)蛋白家族。ABA与受体结合,触发受体发生构象改变,从而使它们与PP2Cs交接,并破坏PP2C-SnRK2相互作用,从PP2Cs介导的抑制作用释放SnRK2s。随后,激活的SnRK2s磷酸化下游靶蛋白,如细胞膜中的离子通道,以及细胞核内的bZIP转录因子。
为了确定ABA信号通路的潜在新组件,该研究小组进行了酵母双杂交筛选,采用SnRK2.6作为诱饵。研究人员发现,1型磷酸酶1(TOPP1)及其调节蛋白,拟南芥(ati-2),可与几个SnRK2s和PYLs相互作用。生物化学和生理学证据表明,TOPP1和AtI-2可协调抑制SnRK2s,反过来,又负调控ABA信号传导通路。topp1和ati-2突变体植物对ABA和盐是高度敏感的,这与它们在ABA信号转导中的功能相一致。此外,转录组分析结果显示,TOPP1和AtI-2共同调节响应ABA的重叠基因群。
总而言之,这项研究确定了ABA信号转导通路新的组成部分——TOPP1和AtI-2,并描述了它们在ABA信号转导中的消极作用。