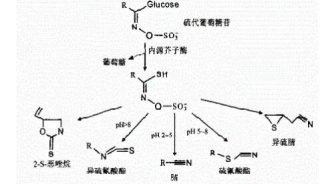
拟南芥中应对硫胁迫的硫的逆向过程
长期以来,植物中的主要(次要)代谢途径一直被认为是将主要代谢产物的前体转化为具有生物活性终产物的一种途径。然而,在环境刺激(如括营养胁迫条件)下,植物组织会出现内源性的终产物降解现象。因此,是否可以将专门的代谢物特别是富含氮和硫的代谢物重新整合到初级代谢中以回收投入其中的资源,对植物来说具有普遍意义。近期,国外研究团队在PNAS上发表了题为“Retrograde sulfur flow from glucosinolates to cysteine in Arabidopsis thaliana”文章,探究了拟南芥中通过将富含硫的代谢物——硫代葡萄糖苷 (GLs)作为硫源将硫基团重新分配到初级代谢物(如拟南芥中的半胱氨酸)的过程。
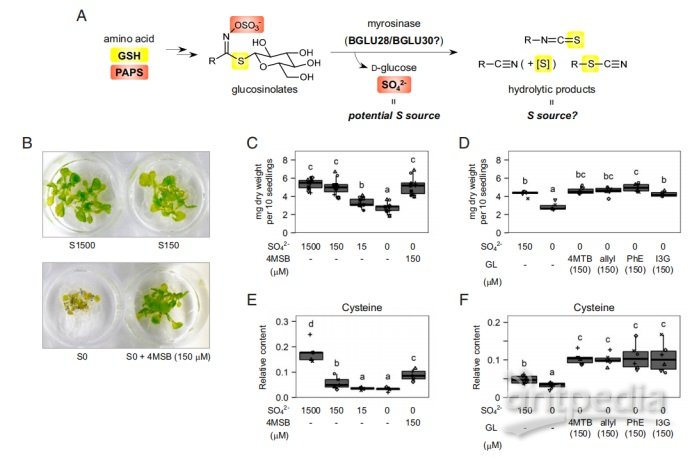
图 1
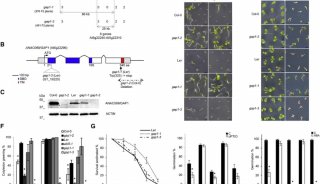
已有的报道显示GL-黑芥子酶(BGLUs)系统具有潜在的硫重新分配作用(图1A)。作者为了探究GLs类代谢物作为硫源的可能性,利用缺硫培养基培养拟南芥,结果显示在缺硫培养基中加入GLs类物质使得其与有硫培养基上生长的拟南芥长势基本一致(图1BCD)。而且使用GLs培养的幼苗其初级硫代谢物(如半胱氨酸、谷胱甘肽)往往高于使用硫酸盐培养的幼苗(图1EF)。这些结果表明GLs可以作为硫源被植物利用。

图 2
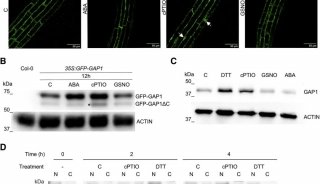
之后,作者使用同位素标记了4-甲基亚磺酰基-正丁基硫代葡萄糖苷(4MSB)中的S(图2A)。当把拟南芥放在用同位素标记S的4MSB作为唯一S源的培养基中培养时,14天后,被同位素S标记的半胱氨酸、甲硫氨酸和谷胱甘肽分别占到了其总量的42%、28%和39%。表明外源施加的4MSB中的S被重新分配到了初级代谢中。

图 3
作者进一步探究了GL水解产物中S所参与到的初级代谢途径。在S1500培养基中培养8天的拟南芥中加入4MSB,其在24小时内完全消耗4MSB并且会伴随着异硫氰酸酯(ITC)和萝卜硫素(SFN)的短暂增加(图3A)。进一步作者使用同位素标记的方法,并且结合各物质的化学结构,探究得到4MSB 水解后的下游途径起始于SFN和GSH的结合,之后逐步裂解SFN-GSH中的Gly和γ-Glu残基,最SFN-Cys环化生成4MSB-NH2和RA(图3B)。

图 4
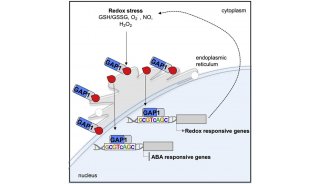
随后作者探究了RA是如何转化成Cys等初级代谢产物,结合已有文献报道作者认为RA可能通过硫氧交换反应释放RA中的硫从而生成原半胱氨酸(pCys),之后通过OXP1生成Cys(图4A)。进一步的实验表明除了高背景水平外,在利用4MSB、RA和pCys处理后,oxp1-1突变体中的pCys积累量要大于野生型中的积累量(图4BCDE)。另外,与pCys一起孵育的oxp1-1突变体蛋白提取物相比于 WT 其Cys的产生率更低,和5OP生成Glu的所得到的结果一致(图4FG)。这些结果表明拟南芥的AtOXP1可以水解pCys,并且RA确实可能通过图4A所示的途径中产生Cys等初级代谢产物。

图 5
除此之外,作者还探究了BGLUs的酶学特性,发现拟南芥的BGLU28蛋白相比于其他GLs对4MTB、苄基硫代葡萄糖苷和PhE的水解率更高(图5A);而拟南芥的TGG1蛋白以相似的速率水解所有类型的GLs(图5B)。为了进一步探究拟南芥BGLU28和BGLU30的功能,作者对bglu28 bglu30双重突变体在不同S条件下培养14天,发现突变体在S15或S0条件下,相比于野生型其生长受到了更严重的抑制(图5CD)。另外,在这两个条件下,野生型植株中的GLs基本都接近检测极限,而突变体中的GLs的积累量明显高于野生型(图5E)。在双重突变体中以4MTB作为硫源时,相比于野生型其生长受到抑制(图1D和5F),并且MTB的积累量也大于野生型(图5G);当用PhE作为硫源时其受到抑制的现象更明显(图5FI)。这些结果表明BGLU28(和 BGLU30)是一种特殊的黑芥子酶,其可以水解特定的GLs,在植物生长的早期阶段其对于硫缺乏的耐受性至关重要。
综上所述,作者发现了植物通过终产物回收利用硫到初级代谢产物的过程,并且用一系列实验证明了这个过程,为植物抵抗营养胁迫提供了一个相对完整的逆向代谢途径。为后续进一步探究植物代谢过程和环境胁迫之间的关系提供了十分重要的理论基础。