用于外源蛋白质生产的细菌表达系统
| 实验方法原理 | |
|---|---|
| 实验步骤 |
一、使用大肠杆菌生产外源蛋白
有越来越多的细菌表达系统可用于外源蛋白的生产。影响选择某个表达系统的因素包括目标蛋白质的天然性质、使用者的经验及产品的预期用途。大肠杆菌是表达外源蛋白最常用的细菌表达系统。经过对大肠杆菌一个多世纪的广泛研究,在调控机制和可能影响表达结果的宿主辅助蛋白的功能方面,研究人员已经获得了大量的信息。此外,实验室和商业蛋白质生产所需的方法和技术有着广泛的来源。对许多蛋白质制备项目来说,这些可用的资源和极低的技术要求使得大肠杆菌成为初步进行蛋白质表达与筛选的首选宿主。因此,我们将用大肠杆菌作为模型,介绍蛋白质生产的具体方法和技术。在介绍使用大肠杆菌进行蛋白质表达的具体技术之后,我们还将探讨其他可用的细菌表达系统。
在此介绍的许多使用大肠杆菌进行蛋白质生产的核心概念和技术可以直接应用于其他细菌表达系统。
二、设计一个细菌表达方案
细菌表达系统的成功运用依赖于表 12.1 中以时间顺序列出的一系列有序的步骤。
蛋白质生产的起始阶段不是开始实验,而是明确对方案的整套要求以及对表达目标的序列特征和生化特性进行分析。这些步骤对方案的最终成功是很关键的,而且有助于对项目成本和成功可能性进行实际评估。为了与实验部分保持连续性,我们对这些步骤进行了简短的介绍,因为在本章的各节中都有对它们更为详细的阐述。
三、方案需求的评估
表达产物的预期用途对于表达系统的选择和决定其优先次序是很重要的。对于某些应用 (如功能解析或药物筛选) 来说,保持蛋白的天然功能和特性是必需的; 但如果是当成免疫原来使用则关系不大。其他需要实际考虑的因素包括蛋白质产量、时间限制或生产成本。明确项目需求对于确保选择合适的表达系统、克隆策略和所需产品的生产策略是至关重要的 (Dieckmanetal.,2006)。
四、目标蛋白质的分析
目标蛋白质的生化和生物学特性对于选择表达策略来说是必须要考虑的,可以通过它们对表达结果和产物的可溶性做出初步预测。通过分析一级序列可以整合一组属性用以预测二级结构、生物学定位 (细胞膜、细胞质、周质腔或胞外)、家族归类 (折叠和结构域) 及推断生化性质 (等电点、无序区域、配基)。目标蛋白质的一些特征,如极可能存在的跨膜螺旋,可能要求必须使用为表达膜蛋白设计的系统,或需要使用克隆单个结构域的策略以表达蛋白质的可溶部分。使用目标蛋白质的特征来指导表达宿主的选择和载体的构建有助于获得成熟的、定位适当的蛋白质。可以使用目标蛋白质或同源蛋白质的实验数据和历史数据对序列信息加以补充,从而有助于表达系统的进一步优化。例如,已知某蛋白质需要某种辅基以保持正确的功目旨,那就必须选择特定的宿主以确保获得具有完整功能的表达产物。一个恰当的例子是: 细胞色素 c 的表达受益于表达辅助蛋白的基因工程大肠杆菌的使用,该辅助蛋白能将血红素共价连接至细胞色素前体的多肽链(Londeretal.,2008)。实际上, 在缺少目标蛋白质或相似蛋白质历史表达数据的情况下,要预测某个蛋白质表达实验的结果是非常困难的。通常的做法是使用数个表达系统, 一开始使用最简单的、性价比最高的表达系统,失败后再使用更复杂的表达系统。
五、克隆
目标序列的克隆有多种选择,通常可将其分为串联系统和并联系统。串联系统使用广泛,为构建表达载体提供了多种生成目标序列/载体匹配末端的方法,如那些利用限制性内切核酸酶生成目标序列/载体匹配末端的方法。该方法的缺点是对每一个目标序列和载体都需要对限制性内切核酸酶的切割策略加以验证。研究者们对平行系统,或者称为普遍克隆技术的兴趣日渐浓厚,该方法可以很容易地将目的片段转移至多种载体和表达系统而无需考虑目的片段的序列(表
12.2)。这种系统的实例包括 Gateway(EspositoetaL,20 〇 9)、Infusion(Zhuetal.,2007)
及不依赖连接的克隆技术(ligationindependentcloning,LIC)(AslanidisanddeJong,1990;Haunetal.,1992)。该方法的优点是能够将一个目的片段克隆至多个不同的载体中,
可以用经济高效的方式同时评估多个表达策略。
LIC 是一种经济高效且特别适合细菌表达系统的技术,因为克隆所用的试剂未ZL化, 且有数家供应商提供相关产品。在 LIC 技术中,特异的核苷酸序列被加至 PCR 引物,这允许克隆任何基因而不需考虑 DNA 的序列。在一种特殊的核苷三磷酸的参与下,通过 T4DNA 聚合酶的处理即可以获得具有匹配末端的载体和 PCR 片段。这一处理可以在载体和 PCR 片段上产生 10~15bp 序列互补的单链突出,这些互补的单链突出退火后具有足够的强度,不需要连接就可用于转化。该方法允许 PCR 引物的设计具有一致性,有方向性,能够获得髙的克隆效率。虽然有商业化的载体可供使用,但该方法已被用于数个大规模的克隆项目,这些项目可以向个别研究者提供载体资源。下面介绍的操作流程是为了使用中西部结构基因组中心(MidwestCenterforStructuralGenomics) 设计的一套载体而开发的 (Eschenfeldtetal.,2009)。只要对添加至扩增引物上的特异序列加以调整,这项技术通常也适用于其他与 LIC 兼容的系统。
六、制备 T4 DNA 聚合酶处理的 DNA 片段
1.在目标序列特异的 PCR 引物上添加合适的 LIC 特异核苷酸序列(如正向引物:TACTTCCAATCCAATGCC; 添加终止密码子的反向引物:TTATCCACTTCCAATGTTA)。
2.该方法可以经放大后使用微孔板进行 (96 个反应),但也可调整至任意数目的反应。混合以下试剂,准备足够用于% 孔板的 LIC 反应混合物。
① 465uL 10 X T4 DNA 聚合酶缓冲液。大部分经销商提供的 T4 DNA 聚合酶缓冲液都可取代本部分介绍的 LIC 反应缓冲液。我们经过比较发现,不同的普通 T4 DNA 聚合酶缓冲液对终产物的克隆效率造成的影响不超过 25%。
② 465uL 分子生物学级的 25 mmol/L dCTP(Promegacat.no.U1221)。
③ 228uL 100 mmol/LDTT(Clithiothreitol) 溶液(Novagencat.no.70099)。
④ 60ul 水。
⑤ 250 单位 T4 DNA 聚合酶 (LIC 品质,约 2.5U/uL,EMD Biosciences/Novagen)。
3.将混合物置于冰上,在使用前加入 T4 DNA 聚合酶。用移液器反复吹打几次以使酶均匀分布于反应混合物中。
4.向聚丙烯 96 孔板中每孔加人 10.4uL LIC 反应混合物。
5.向 LIC 反应混合物中加入 30yLUO?100ng) 纯化的 PCR 片段,用移液器反复吹打几次以混匀。室温孵育 30 min。使用多个片段与载体比例的研究表明,退火反应对于目标 DNA 片段的量的变动有着很大的容忍度。
6.在 75°C 加热块上孵育 20 min,以使 T4DNA 聚合酶失活。
7.在另一块 96 孔板上, 将 1~2 ul T4 聚合酶处理过的 PCRLIC 片段和 4 斗 (20~50ng)T4 聚合酶处理过的 LIC 载体进行退火。
8.在室温下孵育退火反应 5?10 min。
9.使用全部的退火反应体系转化约 50 大肠杆菌感受态细胞, 并挑选出转化的克隆(SambrookandRussell,2001)。
加热后,将 LIC 平板储存于 4°C 冰箱,至需要时再取出。LIC 相容载体的制备与上述过程相似,但需使用碱基互补的 dGTP(Eschenfeldtetal.,2009)。在此阶段,所构建的载体可以通过序列分析进行鉴定,或者在完成表达和可溶性分析之后再进行鉴定。
七、大肠杆菌细胞质中的表达
大部分大肠杆菌表达载体都是设计用于胞内表达的。这些载体经过工程改造,具有不同的筛选标记 (selectablemarker)、细菌启动子、质粒复制原点 (plasmidreplicationorigin)、定位信号 (localizationsignal) 及融合标签 (表 12.2)。我们在此介绍的方法适用于 T7 启动子(Novagen,pET 载体系列),其能够过表达目标蛋白质,表达水平与大肠杆菌中丰度最高的天然蛋白质的水平相当(StudierandMoffatt,1986)。能够表达 T7RNA 聚合酶的大肠杆菌菌株 [如 BL2KDE3)] 对于使用 pET 家族的载体表达蛋白是必需的。
这些菌株的变种能够共表达稀有密码子的 tRNA(Carstens,2003)、蛋白质折叠必需的共辅助因子 (BaneyxandPalumbo,2003) 或是在胞质内有助于二硫键形成和促进重组蛋白活性折叠的那些蛋白质(PrinzetaL,1997)。这里的方案总结了利用 IPTG 和可诱导系统表达目标蛋白质的过程。
八、大肠杆菌细胞质中目标蛋白质的表达
1.接种单克隆于 2 mL 含有合适抗生素的培养基中。所使用试管的体积至少是培养基体积的 5 倍以保证能充分通气。
2.样品在 37℃ 250 r/min 条件下培养至 OD600nm 达到 0.4~0.8。培养液变成云雾状但不要完全变浑浊。通常 BL21(DE3) 菌体达到此状态需要生长 3~4 h。
3.加入 20ul 100 mmol/L IPTG 至每个培养体系(1 mmol/L 的终浓度)。重新将诱导的培养物放回 37°C、250r/min 的细菌培养器,培养 4 h。经过 4 h 的诱导, 培养物会变得完全浑浊。
4.培养物在 37°C 诱导 3?8 h 后应该出现明显的蛋白质表达。取出样品并按下述方法进行分析。
蛋白质表达及可溶性的分析
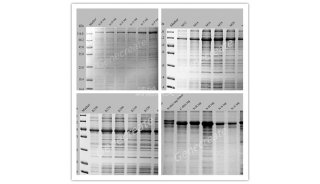
确认目标蛋白质的表达通常涉及对蛋白质表达和蛋白质可溶性的评估,以及对预期蛋白质大小的定性确认。大部分用于外源蛋白质表达的大肠杆菌系统都可以产生出足够水平的目标蛋白质来满足采用变性凝胶电泳的方式分析蛋白质的表达/可溶性
(图 12.1)。这一方法价格低廉,相对简单, 且能很快得到结果。表达水平极低的或可溶性极低的蛋白质可能需要更加灵敏的检测方法,
如免疫印迹法。
九、大肠杆菌中表达的外源蛋白质的分析
1.从每个样品中取出 200uL 诱导物至干净的微量离心管。14000IVmin 离心 Imin。菌体会形成致密的沉淀,同时出现澄清的上清液。剩余的培养物可用于目标蛋白的可溶性质分析,详见本章 10.1 节。
2.倒掉或吸出无用的培养基, 小心保留菌体沉淀。加人 502XSDS 上样染料, 反复吹打几次以重悬菌体。
3.将样品煮沸 5 min,在上样前将样品冷却。对于一个 17 孔的 8 cm X 10 cm 的考马斯亮蓝染色的凝胶,5uL 样品通常就足够了。
蛋白质可溶性的分析
1.通过 3500r/min 离心 10 min, 使剩余的菌体培养物中的菌体形成沉淀。小心倒掉无用的培养基,用纸巾蘸干残留的液体。
2.—80°C 冰冻沉淀。为所有的样品准备足够的裂解缓冲液 [终浓度:300 mmo/L NaCl、50 mmol/L Na2Po4、蛋白酶抑制剂混合物(proteaseinhibitorCocktail,Sigma)、120kU/mL 重组溶菌酶(rLysoZyme,Novagen) 和 25U,mL 核酸酶(Benzonase,Novagen)]。或者,可以用 Bugbuster(Novagen) 或 frPER(Pierce) 代替裂解缓冲液。按照试剂说明,使用适用于 2 mL 培养物的试剂。
3.取出冰冻的样品,稍微解冻。每个样品加入 I80 裂解缓冲液,盖紧盖子后涡旋振荡以重悬沉淀。
4.将样品重新置于一 80°C,5 min, 然后取出并于室温孵育,直到冰完全溶解。再重复冻融一次。样品会变得澄清或是依然保持浑池。
5.3500r/min,离心 10?15 min, 沉淀菌体碎片。
6.从每个样品上部取出 50fxL 上清液, 小心不要带出菌体碎片。
7.向上清液中加人 6 〇 fxL2XSDS 上样染料, 煮沸 5 min。用变性凝胶电泳分析样品。对于一个 17 孔的 8 cmX10 cm 的考马斯亮蓝染色的凝胶,5 卟样品通常就足够了。
8.(可选) 为了检测菌体裂解物中的不溶部分,除去步骤 7 菌体裂解碎片中所有剩余的上清液。小心不要扰动或移走沉淀。
9.向沉淀中加入 300juLIXSDS 上样染料,盖紧盖子后涡旋振荡至沉淀完全重新悬浮。
10.煮沸样品 5 min, 在上样至 SDS-PAGE 凝胶前稍加冷却。对于一个 17 孔的 8 cmX10 cm 的考马斯亮蓝染色的凝胶,3 样品通常就足以观察表达情况。
蛋白质表达结果的分析
在评估表达和可溶性时使用同样的培养系统可以弥补表达水平的差异。在大肠杆菌中蛋白质表达率通常可以超过 80%,但蛋白质的可溶性是该系统的主要限制因素,它与目标蛋白质本身有关(图 12.1)。经过 SDS~PAGE 分析,如果在正确的分子质量位置未检测到目标蛋白质的染色条带,那就可以认为其「不表达」或是「不可溶」。反之,则认为其可以「表达」或「可溶」。这属于定性的评估,必须再进行序列分析和/或质谱分析以确认^肽的身份。我们采用了相对密度排序法,即将目标蛋白质的染色密度与大肠杆菌中天然蛋白质的通常染色密度相比。目的条带可见,但其染色密度比大部分大肠杆菌蛋白质条带的染色密度低,则记为 1 级水平或是低表达/低可溶性。2 级和 3 级水平(中等或高表达/高可溶性) 分别表示染色强度与大肠杆菌中高表达的蛋白质染色强度相当或更高。
蛋白质表达的自诱导方法
IPTG 诱导的一个缺点是需要监测细菌的生长以获得最佳的诱导条件。这一缺点在检测大批量克隆的表达时变得尤为突出。在这种情况下,可以观察到生长速率的不同,从而很难获得所有表达克隆的最佳诱导条件。自诱导系统为我们提供了克服这些困难的方法, 并且简化了表达方案(Studier,2005)。目前已报道了数种自诱导系统, 它们可以利用 pET 和其他 IPTG 诱导的细菌表达系统来提供高水平的蛋白质表达 (Blotnmeletal.,2007)。
十、使用自诱导培养基方案的小量表达培养
十一、蛋白质的周质表达
细菌蛋白质组中有 8%?12% 的蛋白质不定位于胞质中。针对这部分蛋白质的外源表达策略包括: 扩增编码区的非信号肽部分,利用胞质表达载体或周质表达载体,通过标准的大肠杆菌克隆和表达流程进行操作, 从而得到与胞质蛋白相似的表达结果。该方法也成功用于某些含有二硫键的真核蛋白。通过在目标蛋白质 N 端添加合适的定位信号 (如大肠杆菌的 PelB 信号序列),就可将蛋白质定位于大肠杆菌的周质空间。由于周质只占菌体总体积的 20%?40%,总的来说,周质表达的表达量一般低于胞质表达的。直接比较克隆至胞质表达载体的目标蛋白质 (图 12.2, 目标蛋白质 4?6) 与克隆至周质表达载体的目标蛋白质 (图 12.2, 目标蛋白质 10?12) 的表达水平,就可得出此结论。
十二、大肠杆菌周质腔中目标蛋白质的表达
1.接种单克隆于 2 mL 含有合适抗生素的培养基。所用试管的容纳体积至少是培养液体积的 5 倍,以保证充分的通气。
2.30°C、250r/min 条件下培养至 OD68jnm 达到〇.4?0.8。培养液应该变成云雾状但不要完全变浑浊。通常 BL2KDE3) 菌体达到此状态需要生长 3?5 h。
3.降低培养容器温度至 19°C,孵育至少 30 min 以使菌体达到平衡。培养物的 OD 值在温度转换期间会继续增加。
4.加人 20 斗 100 mmol/LIPTG 至每个培养体系 (终浓度为 Immol/L)。将培养体系放回培养容器,19°C、250r/min 过夜培养。经过诱导过夜,培养液会完全变浑浊。
5.培养物在 19°C 诱导 4 h 后就会有少量表达,12~16 h 后就会有明显的表达。
十三、小规模渗透休克法
如果需要确认一个目标蛋白质定位于周质中,那么渗透休克法可用于分析与胞质组分分开的周质组分。有许多从细菌周质中释放蛋白质的技术可供使用。加入三氯甲烷
(chloroform)(Amesetal.,1984) 或多黏菌素 B(polymyxinB)(DixonandChopra,1986)
可以从周质中释放蛋白质。下面介绍的方法成本低廉,使用的是常用的实验室试剂
(NeuandHeppeU1965;NossalandHeppeU1966)。将溶菌酶(lysozyme) 加人渗透休克 SETC
蔗糖、EDTA、Tris-HCl) 缓冲液中可以除去更多的菌体外壁和组分
(BirdsellandCota-Robles,1967;MalamyandHorecker,1964);
这一步产生的原生质体(spheroplast) 非常容易裂解,因此胞质蛋白可能会污染周质组分。对于易裂解的菌体,向冷水休克液中加入〇.5
mmol/L 的 MgCl2 可以稳定原生质体
(NeuandChou,1967)。我们介绍的方法对大部分耙向周质的蛋白质都适用,但对有些靶向周质的蛋白质可能需要优化。比较图 12.3 中
SET 和水中目标蛋白质的量,可说明对不同蛋白质来说抽提结果是不同的。不过, 相比于背景宿主蛋白,可溶的外源蛋白富集在休克组分中(图
12.3)。该方法可以放大,作为第一个纯化步骤以减少背景蛋白质和菌体碎片。
十四、用于外源蛋白质生产的其他细菌系统
有许多不同的细菌表达系统可用于表达外源蛋白。许多时候,细菌表达系统的选择通常是由目标蛋白质的特性决定的。尽管大肠杆菌可用于多血红素细胞色素 (nmltihemecytochromes) 的外源表达,但通常会选择另一种宿主来表达含多个血红素基团的细胞色素复合物(complexcytochrome)。希瓦氏菌(S/iewaneWaonei<ie7m\s) 是一种革兰氏阴性菌,通常用于生产这种特殊的蛋白质。该生物的基因组能编码大量预测的细胞色素 c 基因,而且含有能促进细胞色素 c 被正确加工成为成熟蛋白的辅助蛋白(TakayamaandAkutsu,2007)。一个经过工程改造的突光假单胞菌(_Psewc?omonas/ZM 〇?-e?:e?w) 菌株 (Pfenex,DOWChemicalCa) 已经被开发为外源蛋白表达系统,用于大批量发酵和目标蛋白质的高通量小规模筛选。这一经过修饰的革兰氏阴性菌具有多种代谢功能和微生物分泌系统,从而可以使表达产物位于胞质、周质或胞外空间。
利用非致病性革兰氏阳性杆菌表达蛋白质为我们提供了革兰氏阴性宿主菌之外的另一个选择 (Schmidt,2004)。这些生物含有天然高效的分泌系统,可以直接将蛋白质表达于培养液上清液中,从而可以提高产量,同时便于纯化。最常用的菌株是工程改造的枯草芽孢杆菌 (BadWws,其缺乏胞外脂肪水解酶 (lipolyticenzyme) 和蛋白酶的基因,从而提高了外源蛋白的稳定性。巨大芽抱杆菌 (BaciZZwsOTegaferiwm) 有数个大质粒,它以能够稳定复制和保持这些质粒而闻名。其表达菌株通常具有较低的内在蛋白酶活性,还有一些特点组合在一起能够获得蛋白质的稳定高产。
膜蛋白在基因组编码的蛋白质中占有相当的比例,但对于蛋白质表达来说仍然是个挑战。除了大肠杆菌(NeophytouetaL,2007;ShawandMiroux,2003),研究者们还开发了数个其他的膜蛋白表达系统来表达完整的膜蛋白。革兰氏阳性的乳酸菌—乳酸乳球菌(LartococcwsZartt’s) 生长密度高,适于大规模的膜蛋白超量生产(Kunjietal.,2003)。有数个营养缺陷型菌株和多环乳酸链球菌素(polycyclicpeptidenisin) 调控的可诱导表达系统可以使用。用于膜蛋白鉴定的功能筛选可使用全菌进行,因为配基可以直接作用于表达在细胞膜上的膜蛋白。
经过工程改造,光合细菌红细菌属已用于全长膜蛋白的外源表达 (Laibleetal.,2004)。该系统可以通过在表达外源膜蛋白的同时合成新膜的方式来将外源膜蛋白整合于其自身的内源膜蛋白系统中, 这一点是很独特的。
这些新的细胞膜形成使细胞质膜的内凹,允许外源膜蛋白整合。红细菌属膜蛋白表达系统非常强有力的一点是在各种情况下表达的蛋白质都几乎等量地定位于膜。膜蛋白的表达和纯化是有难度的,而且用外源表达系统生产的蛋白质的功能必须经过仔细的验证。
十五、其他载体和诱导条件
除了使用不同的细菌宿主外,外源表达系统的优化还可通过控制载体和诱导条件来实现。其中一个已被证明成功用于蛋白质可溶性表达的此类方法使用了 pCold 载体的冷休克表达系统(Qingetal.,2004)。该系统利用了大肠杆菌细胞的冷休克反应以抑制内源蛋白的产生,同时又增强了目标蛋白质的表达。研究表明,当截短的冷休克蛋白 (CSpA)mRNA 在冷休克的条件 (从 WC 降温到 15°C 并保持 36 h) 下表达时,多聚核糖体就会被占据用来翻译截短的 c 分 A 基因,而不能形成 HI 型核糖体去翻译非冷休克蛋白 (non-cold-shockprotein)(JiangetaL,1996)。结果是,只有在 mRNA 中的基因才会被翻译,而内源蛋白的表达则受到抑制。在 pCold 质粒中启动子之后有一个克隆区,用于插入目的基因,使用 1mmol/L IPTG 可以阻遏目标蛋白质的表达,15℃ 冷休克可以诱导目标基因的过表达,由于宿主背景蛋白的表达极低,该技术表达的外源蛋白通常可以不用纯化,这大大消减了生产成本。
展开 |