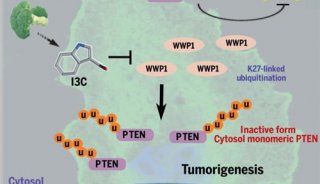
清华柴继杰教授解析自交不亲和反应分子机制
生物通报道:11月8日,国际学术期刊《Cell Research》在线刊登了清华大学、中国人民大学和康奈尔大学的一项重要研究成果,题为“Structural basis for specific self-incompatibility response in Brassica”。这项研究首次阐明了自交不亲和性(SI)在十字花科家族的分子机制。
这项研究的通讯作者是清华大学博导、杰青、长江学者、973首席科学家柴继杰教授。其1997年于协和医科大学药物研究所获得博士学位;1997-2004年先后在中科院生物物理研究所和普林斯顿大学从事博士后研究。柴继杰教授带领的课题组关注并研究在生物学及药学应用中的重要大分子结构与功能。主要通过蛋白晶体衍射的方法及一些细胞生物学、生物化学等手段阐述这些生物大分子在结构和功能上的联系。相关阅读:清华北大合作发表多篇文章解析CLE家族小肽;凋亡复合体和炎性体:caspase活化的保守装;清华、北大合作发表Cell Rep文章解析激素受体。
在开花植物中,自交不亲和性(SI)是避免自交和近交、从而保持其遗传多样性的一种普遍机制。在十字花科植物中,SI是由一个高度多态性的基因位点的变异单倍型介导的,称为S位点,通常包含三个高度多态性的基因:SRK、SCR和SLG。遗传和生化研究将SRK和SCR确定为SI特异性的唯一决定因素,将SRK确定为SCR的受体,可使柱头区分SI反应中的“自我”和“非自我”花粉。由于SCR蛋白将会结合并激活编码在同一S位点单倍型中的SRK变体,SRK和SCR蛋白必须共同进化来维持它们的相互作用。通过SRK胞外结构域(eSRK)对“自我”SCR的这种高度特异识别,可诱导SRK激酶的激活,从而触发信号级联反应来抑制“自我”花粉。SCR增强的SRK同源二聚化对于这种信号的开始,是很重要的。
SRK属于大的植物受体家族,受体激酶(RKs),其中包括超过十个亚科,在不同的生物过程中起着重要的作用。SRK是S域PLK(SD-RLK)亚科的原型成员,大约40个编码在拟南芥基因组中。eSRK的特征是两个连续的N端凝集素样结构域,随后是含有12个保守的半胱氨酸残基的区域。结构建模预测六个N-末端半胱氨酸包含有一个EGF样结构域和六个C-末端半胱氨酸。eSRK序列的比较,确定了几个超变量(hv)区域,指定为hvI、hvII和hvIII,据预测,这对于SI的特异性是非常重要的。一致的是,对于少量变异研究表明,来自hvI和hvII区域的残基,是“自我”SCR识别所需要的。但这些区域是否是所有SCRs被它们的同族SRKs识别所需要的,仍然是未知的。在SCR变异体的情况下——它们比SRKs更多态,一项NMR研究表明,这些50个氨基酸的小蛋白,通常含有八个保守的半胱氨酸,可能都有一个类似于防御的结构。在两种甘蓝SCR变种(SCR6和SCR13)中开展的一项研究表明,位于第五个和第六个保守半胱氨酸之间的四个连续的氨基酸残基,对于SCR13被SRK13的特异性识别至关重要,但不能用于SCR6被SRK6的识别。因此,尚未出现一般的规则来预测,受体和配体中的哪些残基决定着它们的特异性相互作用。
为了填补由于“缺乏SRK识别SCR的根本结构机制”所造成的缺口,该研究小组解析了eSRK9-SCR9复合物的晶体结构,这原来是一个2:2 eSRK:SCR异四聚体,形状像字母“A”。 eSRK9的所有三个hv区域,介导SCR9的特异性识别。有趣的是,SCR9可通过与来自一个eSRK9单体的一半hvII,以及来自第二个eSRK9单体的另一半hvII相互作用,诱导eSRK9的二聚化,而不涉及两个SCR9分子之间的相互作用。总而言之,这些研究结果首次阐明了SI在十字花科家族的分子机制。