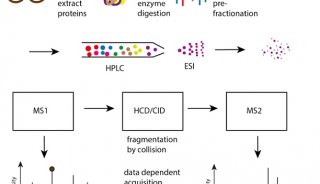
| 实验步骤 | 第一阶段 用荧光染料标记蛋白质
材料
缓冲液和溶液
将储备溶液稀释到适当的浓度。
用 NaOH 调节的 N-二羟乙基甘氨酸(0.1mol/L,PH7.5)
用 NaOH 调节的 N-二羟乙基甘氨酸(1mol/L,pH9.0~9.5)
消化缓冲液
20 mmol/L 磷酸钠
10 mmol/LEDTA
20 mmol/L 半胱氨酸/HCl 缓冲液(pH7.0)
N,N-二甲基甲酰胺(DMF)
在存储容器中加入约 1/3 体积的吸湿性树脂用于干燥 DMF。例如,可以始终将 AG501-X8 混合床树脂 (Bio-Rad) 放在 DMF 原料瓶中。如果对 DMF 的完整性有任何疑问时,则可以另外加入一些新鲜树脂,或者用新鲜树脂替换老树脂。这样的操作是考虑到会出现标记蛋白质能否达到期望比例的问题,因为标记反应时水的存在(可能来源于 DMF) 会竞争游离染料。
含有 10 mmol/LEDTA(pH7.0) 的磷酸钠(20 mmol/L)
磷酸盐缓冲溶液(PBS)
TE(pH7.0)
酶和缓冲液
固定在琼脂糖顆粒上的木瓜蛋白酶(分散在消化缓冲液中的 50% 混悬液;Sigma)
标记化合物
Cy3 和 Cy5 OSu 单功能硫代吲哚花菁琥珀酰亚胺酯(Amersham)
硫代吲哚花菁(cy) 的琥珀酰亚胺酯染料共价结合到游离氨基(赖氨酸侧链α-氨基末端或 e-氨基)。Cy 染料以冻干态使用,所有情況下保证其处于干燥状态以避免与水反应。由于批与批之间可能存在差异,每一瓶中染料的量必须单独测定。在这里所举的例子中,用俄勒冈绿的琥珀酰亚胺酯(分子探针)来标记 MC5 PKCα抗体。
抗体
待标记的纯化抗体,如 MC5(单克隆)或 T250(多克隆),或蛋白质标记反应必须在没有氨基的缓冲液中进行。许多商用抗体制品中添加了含有可能竞争 Cy 染料的游离氨基添加剂(如牛血清白蛋白或明胶)。也有许多供应商可以根据要求提供不含这些化合物的抗体制品,这种情况下可以要求提供溶于磷酸盐缓冲溶液中、浓度为 lmg/ml 的样品。
凝胶
SDS-聚丙烯酰胺凝胶
制备 SDS-聚丙烯酰胺凝胶分离蛋白时,请参考附录 8
专用装置
Centricon 浓缩器:YM10,YM30,YM100(分别切割 10、30、100kDa 大小的分子;Amicon)
凝胶过滤/分子大小排阻色谱柱(预装一次性 Econo-Pac 10DG;Bio-Rad)
蛋白 A-琼脂糖凝胶柱(Econo-Pac 的 2 ml 柱;Bio-Rad)
用装料体积 20 倍的 20 mmol/L 磷酸钠冲洗柱来使柱平衡。推荐用蛋白 G 来纯化小鼠抗体。
UV/可见吸收分光光度计
附加试剂
本方案中的第 7 步需要测定蛋白质浓度的试剂(如 Bradford 分析试剂)(Spector et al.1998,第 56 章)。
方法
Fab 片段的制备与纯化
1. 采用像 Centricon YM100 的一次性浓缩器,以离心的方法使抗体在含 10 mmol/L EDTA 的 20 mmol/L 磷酸钠溶液(pH7.0) 中的浓度浓缩至 15~20 mg/ml。
2. 向 250ul 浓缩的抗体制品(约 4~5 mg) 中加入 500ul 消化缓冲液和 500ul 固定在琼脂糖顆粒上的 50% 木瓜蛋白酶混悬液(分散在消化缓冲液中)。37°C 振摇(振荡孵化器中,300~400r/min)6~10 h 进行消化。
重要:消化过度(如超过 16 h) 将导致 Fab 片段裂解成较小的多肽。因此建议进行预试验,毎隔1h 取少量的消化液(约 5ul) 采用还原性 SDS 聚丙烯酰胺凝胶电泳进行分桁,在还原条件下,lgG 的重链和轻链会像 50kDa 和 25kDa 的多肽一样迁移。经木瓜蛋白酶消化的抗体按约 25kDa 大小的分子迁移。
3. 采用蛋白质 A-琼脂糖凝胶柱从 Fc 部分和未经消化的抗体中纯化 Fab 片段。将消化混合物加到平衡过的柱上,并按每份 1 ml 立即开始收集流过液部分。
蛋白 A 将与抗体的 Fc 部分结合,因此可以从消化混合物中除去全抗体和 Fc 部分,获得的流过液就仅仅含有 Fab 片段。注意!IgG 分子的许多亚类不与蛋白 A 结合。当需要制备小鼠单克隆抗体的 Fab 片段时,使用蛋白 G。
4. 为了鉴定哪些流过液部分中含有纯化的 Fab 片段,测定每一部分的 OD280, 收集并合并 Fab 浓度最高的三个或四个组分。
一般来讲,可以在大约 3~4 个组分中发现 Fab 片段。
5. 使用 Centricon YM100 柱(截流值 10kDa;Amicon) 离心将合并的组分浓缩至体积 0.5 ml 以下。
6. 将样品加到 PBS 平衡的低分子质量(6kDa 以下)凝胶滤过层析柱上,用 PBS 洗脱 Fab 片段(按照产品说明进行)。
为了标记 Fab 片段,必须将它们交换到不含游离氨基(如消化液中必须除去游离半胱氨酸)的缓冲液中。
7. 洗脱后,采用 Centricon 浓缩器使 Fab 片段浓缩后,测定蛋白质浓度。加入 1/10 体积的 1mol/L N-二羟乙基甘氨酸(pH9.0)
采用 Bradford 分析法测定 IgG 浓度,以牛血淸白蛋白作为标准品,并乘以系数 2。
浓缩的 Fab 制品现在就可以准备进行荧光硫代吲哚花菁染料标记。
染料制备
8. 将各瓶冻干染料中的内容物重新混悬于 20ul 的干 N,N-二甲基甲酰胺中,得到浓度约 10 mmol/L 的 Cy 溶液。
9. 将 1ul 染料混合物用 PBS 按 1:10000 稀释,并由可见吸收峰计算染料浓度。
Cy3 和 Cy5 在 554nm 和 650nm 的吸光系数分别为 150000L•nmol-1 和 250000L•nmol-1。
染料结合
10. 标记蛋白质时,将其混悬于不含游离氨基的缓冲液中。如果蛋白质溶液不是在一种适当的缓冲液中,则按上述第 6~7 步进行操作。
蛋白质如果因稳定性的考虑必须采用持定的缓冲系统,则凝胶过滤系统也采用该缓冲液平衡。注意缓冲成分中不应含有游离氨基或抑制标记反应的成分。值得注意的是,像二硫苏糖醇、β-疏基乙醇等还原剂对标记反应有干扰。然而,如果为了保持生理/蛋白质功能而必须添加还原剂时,β-疏基乙醇因为干扰较低,因此是首选的还原剂。在 0.1mol/L N-二羟乙基甘氨酸(PH9.0) 中进行标记以前,T(P)250 和 MC5 均需置换到 PBS 中。
建议在标记反应过程中保持碱性 pH 条作(同时保持蛋白质的完整性),以便使赖氨酸的ε-氨基增加质子解离性并因而能有效地结合。
11. 蛋白质样品(抗体、Fab 片段或蛋白质)与 10~40 倍过量摩尔数的 Cy3 在室温反应 30 min(按第 8~9 步所描述的进行)。将染料非常缓慢地加入到用移液管尖头搅拌的溶液中(小心操作以避免染料直接与微离心管接触)。
为了避免蛋白质因为 DMF 而变性,所加人的 Cy3/DMF 体积必须不超过总体积的 10%。
最佳的过量摩尔数以及反应时间都是根据经验来确定的。标记的目标是能使毎一个蛋白质分子与 1~3 个染料分子结合(对于标记比例标准的讨论,请参阅本方案的导言部分)。对于不同的蛋白质或抗体,其标记水平有很大的变化。如果最终的标记率太低,可能可以重新标记抗体。像 T(p)250 和 MC5 等抗体,已经成功地用超过 40 倍过量摩尔数的染料进行标记。
12. 加入最终浓度为10mmol/L 的含有游离氨基的缓冲液,如 Tris 等来终止标记反应。
13. 采用凝胶过滤柱系统(10DG,Bic-Rad) 来交换缓冲液以除去多余的未反应染料,并将蛋由质洗脱至 PBS 中。
a. 用三倍量填料体积的 PBS 或其他选定的缓冲液(约 30 ml) 平衡柱系统。
b. 为了达到最大分离,将标记反应的混合物直接加到树脂上,并注意使它的体积越小越好。
c. 用 2.5 ml 缓冲液(上空体积约为 3.3 ml) 清洗柱并废弃洗脱液。
d. 另外加入 2 ml 缓冲液并收集可见蛋白质组分,将剩余物丢弃。标记产物将跑在前沿,并可以肉眼观察到,而游离染料从柱上洗脱较慢。
14. 采用 Centricon 浓缩器(可以截流适当大小的分子)再次对标记蛋白质进行浓缩。
15. 采用 SDS 聚丙烯酰胺凝胶电泳分析标记产物,以便验证蛋白质是否成功地被标记。产物经电泳分离后,直接用紫外透照仪(302nm) 检査凝胶上的可见标记产物。
荧光带迁移至蛋白质的预期分于质量部位。在电泳迁移的前沿没有游离染料的荧光;
对于大多数染料,在 302nm 的紫外激发是较好的选择。如果对凝胶照射时不能看到带,则可以考虑采用更短波长的激发光。
16. 标记反应以后,采用下述公式计算标记率:
Aλ X M / [ ( A280-ƒ X Aλ ) X ελ ]
式中 Aλ是染料在其最大吸收波长λ处的吸收度,A280 是蛋白质在 280nm 的吸收度,M 是以 kDa 为单位的蛋白质分子质量,ελ是染料在波长 λ 处的摩尔消光系数,以 L•mmol-1•cm-1 为单位。公式以染料在 280nm 处的吸收进行校正。系数ƒ是染料在 280nm 和最大可见吸收波校λ的吸收度比值。例如,Cy3 标记抗体的标记率公式就变为:
A554 X 170 / [ ( A280 - 0.05 X A554 ) X 150 ]
为了验证染料的结合是否会影响 Fab, 整个抗体或者蛋白质的生理功能,将标记产物与相应的未标记成分进行持异性活性比较。特异性活性包括催化活性或蛋白质结合活性。这些蛋白质结合分析可以采用琼脂糖固定的肽配体或磷酸酪氨酸颗粒来进行。这种结合活性测试的标准探作规程可以参阅《抗体技术实验指南 (Using Antibodies)》(Harlow and Lane1999)。(该书中译版已由科学出版社 2002 年 9 月出版。------编者注)
第二阶段 FLIM.FRET 分析的细胞制备
材料
缓冲液和溶液
将贮存溶液稀释到适当的浓度。
明胶溶液(0.1%)
可选,请参见第 1 步。
将 0.5 g 明胶(Porcine,Sigma) 溶于 500 ml 1X 麻酸盐缓冲液液中。高压灭菌。
磷酸盐缓冲溶液(PBS)
聚-L-赖氨酸
可选,请参见第 1 步。
可以从 Sigma 购得 0.1%(m/V) 水溶液,使用前用水按 1:10 稀释。
核酸和寡核苷酸
载有探针的质粒 DNA
编码了目标蛋白质的 GFP 融合载体(CLONTECH)。
培养基
CO2-非依赖性成像培养基
这种低背景的荧光培养基可以从 Life Technologies 购得,或者采用经 Dulbecco 修改 Esgle's 的培养基的标准组成中除去下列成分以后的培养基:pH 标记物酚红、青霉素、链霉素、叶酸以及维生素 B2。使用前采用以 NaOH 调至 pH7.4 的无菌 HEPES(终浓度为 50 mmol/L) 来补充培养基。
成像培养綦必须不含有自动荧光成分。
专用装置
组织培养板(6 孔或 12 孔)(Nunc), 或者
玻璃底组织培养皿(35 mm,MatTek Corporation),或者
腔室载玻片或腔室盖破片(Labtek,Nunc)。
附加试剂
本方案中的第 2 步需要用第 16 章中介绍的转染方法所需的试剂。
细胞和组织
进行转染的细胞株(贴壁或悬浮培养)
除了考虑细胞株是否适合所研究的系统以外,选择细胞类型惟一需要考虑的就是通过—些方法可以方便地将它们固定(请参见第 1 步说明)。
方法
1. 将细胞移植到适当的表面进行显微观察:
对于活细胞制品:将细胞铺到玻璃底平皿或腔室盖玻片中。
对于转染后固定的细胞制品:将细胞移植到置于 6 孔或 12 孔组织培养皿中的玻璃盖玻片上。
容器的最终选择取决于细抱在转染后是否需要微注射;如果需要. 则应根据针头操作需要来考虑器皿的形状和剩余空间。
对于悬浮细胞,固化柯以更方便地使用像明胶(0.1%,m/V,PBS; 高压灭菌)或聚 L-赖 氨酸(0.01% 水溶液,m/V) 等培养基。两种情况下, 均采用所选择的培养基覆盖小孔、盖片或者盖玻片约 30 min 来对上述材料表面进行包覆,再吸出过剩的培养基,然后,将细胞直接移植到培养基裏盖的表面。在转染操作过程中,细胞也可以保持在混悬液中,到成像前再立即固化细胞一旦固化后,就可以进行微注射了。
将细胞按-定的密度涂板,使约 40% 的细胞在第二天可以融合。
2. 采用第 16 章中所介绍的任意一种转染方法,用编码了 GFP 标记的目标蛋白的质粒转染细胞。
3. 转染细胞在适当条件下孵化 16~24 h, 以使细胞表达目标蛋白。
在有些情况下,必须梢确控制蛋白质表达水平。此时,建议在进行 FLIM 測定前先优化转染效率以及表达周期,可以采用调整的增强子来促进蛋白质的表达(请参见第 17 章),或者采用核微注射来替代转染(请参见本方案结束处的“替代方案,活细胞的微注射”)。
4. 表达了目标蛋白质的细胞的鉴别。
5.(可选)如果实验设计允许, 选择第 6 步后面的替代方案,即微注射 (用于活细胞)或者固定和染色 (用于固定细胞),中的一种方法来导入另一探针 (如: 标记蛋白)。
6. 在进行显微实验以前,立即用 CO2 非依赖性成像培养基(市售培养基或按材料项介绍的方法调整的经 Dulbecco 修改的 Eagle’s 培养基) 替换培养基。
在实验设计中,如果在有关的处理前需要将细胞血清饥饿,则在成像前按所介绍的时间段让细跑饥饿。但是, 如果细胞在缺乏血浆时会发生凋亡的话,就应在饥饿期间将细胞置于 0.5% 的胎牛血淸中,然后在成像前立即用无血淸培养基替换。




第三阶段 FLIM·FRET 测量
材料
专用装置
放大器(ENI403LA 或 Intra-actionPA-4)
氩激光(Coherent,Innova70C)
宽带绝缘反射镜(2 个;Newport Corporation)
检测器
MCP,HamamatsuC5825 或 LaVision Picostat HR
CCD, 配备 Kodak KAF 1400 芯片的 Photometries Quamix
滤光器
GFP,OG(Q495LP,HQ5lO/20;Chroma)
Cy3(HQ545/30,Q565LP,HQ610/75;Chroma)
单面镀银反射镜(Zeiss)
曲镜座(2 个),透镜支架(2 个),标竿(5 个),以及后支架(5 个)(Newport)
倒置显微镜(Zeiss 135TV)
IPLab Spectrum(Signal Analytics)
可变光圈(Comar)
透镜(分别为焦距 12 和 7.6 cm、焦距 2.5 和 3.8 cm;Newport)
水银灯(100 W Zeiss;HBO 100 AttoArc)
Multimode fiber step-indexed 1-mm core(步长指数 1 mm 内核的多态纤维)
(Newport)
油接物镜(100X/1.4NA;ZeissFluar)
光学操作台(2X1m;TMC)
Power meter 和 head (Ophir,Nova Display 和 2A-SH)
配备 PCI-GPIB 卡(National Instruments) 的 Power PC(Macintosh)
快门(高速)Vincent Associates,UniblitzVS25)
快门启动器(Vincent Associates,UniblitzVS25)
驻波声-光调制器(acousto-optic modulator,AOM)(Intra-action,80MHz)
倍频合成器(IFR2023)
可变密度滤光器滑轮(Laser Components)
水循环冷却装置(Grant)
细胞和组织
按第二阶段所介绍的方法制备表达了蛋白质探针的细胞(活的或固定的)
方法
1. 调节波长选择棱镜,选定 488nm 的氩激光为激发波长。采用激光背面的控制旋钮微量调整高反射镜面的位置来优化输出功率。
2. 设定频率合成器来启动 AOM 至共振频率约为 40MHz(对于这里所介绍的实验,采用的启动频率为 40.112MHz)。这使得激光束以两倍于驱动频率(80.224MHz) 的强度振动。
3. 通过调节入射角的角度和用光度计监测非衍射零级光的强度来优化 AOM 的衍射以及调制强度。在优化的衍射角度(达到相应的最大衍射)时零级光的输出功率最小。
4. 开启 MCP 和 CCD。将光阴极电压的基底值设为-2V, 并调整增益以匹配 CCD 的满动力范围。这取决于样品的荧光强度,并需要凭经验来确定。最理想的是,使增益值尽可能小以降低噪声。通常将 Hamamatsu C5825 的增益值设定为 1, 而 Photometrics Quantix CCD 的增益值设定为 3。将 CCD 的示值读数设定为 2X2 栅格。
5. 将主频率合成器驱动 MCP 的频率设定为驱动 AOM 频率的两倍(对于上面所举的例子即为 80.224MHz)。
6. 选择最合适的物镜来进行实验。在本例中使用的是 Zeiss Fluar 100x/1.4NA 油物镜。
7. 为了得到零级寿命参照图像,从强扫描开始每相隔 22.5°记录整个一周 16 张相位依赖性图像(例如:将一小片铝箔置于盖玻片或玻璃底平皿的成像表面)。
a. 将荧光滤器部件改为单面镀银反射镜,并用可变强度滤光滑轮将外来光的强度调至最小值。将聚焦点调整至铝箔表面。
注意:当设置铝箔成像时,在将外来光源的强度调至最小值以前,千万注意不要直接用肉眼看显微镜。
b. 照一张曝光时间约为 100ms 的铝箔图像。这张图像是用来选择 ROI 并估计所需的最佳曝光时间的。因为此时主频率合成器的相位可能不是最大的,为了避免检測器的饱和,应该选择可以生成约 1000 个计数的曝光时间。
c. 每相隔 22.5°记录整个一周 16 张相图,测定图像系列中达到最大强度时的相位,并在主频率合成器上重新将该相位设为 0 度。
d. 记录铝箔另一周十六张相图并记为零级寿命参照图像。
当进行寿命成像时,我们建议随时监测相稳定性。我们的装置一小时内相位可以稳定在 0.3°以内。对于我们的装置,每小时记录并保存一组参照铝箔像序列。
8. 将外来激发光源保存为最大(利用可变强度滤光滑轮);这样系统就已经准备好可以采集细胞的图像资料。
9. 用 GFP 滤光器设置(分光器:Q495LP, 激发装置:HQ510/20) 获取一张供体图像。曝光时间设定为能达到 75% 的 CCD 动态范围(对于 12 位 CCD 为 3000 个计数)。在图像中选择感兴趣的区域来測定荧光寿命。
10. 制备供体荧光寿命图谱:
a. 获取两个邻近的、一个正相另一个反相循环的 16 张相位依赖性图像(间隔 45°相位)系列来校正相淬灭。按第 9 步的方法优化每一张相图的曝光时间设置。
b. 在没有样品照射的情况下获取另一张图像。将这张去除背景的图像从系列的所有相图中提取出来,然后将正相和反相循环中每一对相同相位的图像进行一级加和来修正供体的淬灭。
c. 由这些资料以及零级寿命参照图像(铝箔),按照本方案导言部分关于图像操作的讨论中所介绍的方法来计算供体荧光寿命图谱。
11. 记录受体的图像,然后将照明光源改为 100W 水银灯(Zeiss Attoarc) 来淬灭受体。将 Cy3 滤光部件(激发装置:HQ545/30; 分光器:Q565LP; 发射装置:HQ610/75chroma) 移到检测通路中。将曝光时间优化到可以占满 CCD 整个动态范围后,获取受体图像。在不能检测到 Cy3 荧光的条件下,关闭检測器部件,照射受体。
重要,当进行受体的光漂白实验时,明确光漂白以后,在供体通道内的受体的光产物没有荧光这一点是极为重要的。





 |
|---|