分享至
分享至



综述 | Cell子刊:胆汁酸-微生物互作及对宿主代谢的影响

肠道微生物群被认为是代谢“器官”,不仅有助于从摄入的食物中获取营养和能量,还产生许多代谢物,通过其同源受体发出信号来调节宿主代谢。胆汁酸就是一类这样的代谢物,由肝脏中产生的胆固醇通过肠道微生物群在肠中代谢形成。胆汁酸反过来又可以通过激活小肠中的先天免疫基因直接或间接地调节肠道微生物组成。因此,宿主代谢可以通过胆汁酸的微生物修饰而受到影响,胆汁酸通过胆汁酸受体改变信号传导,也改变微生物群组成。
论文ID
原名:Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism
译名:胆汁酸-微生物互作及对宿主代谢的影响
期刊:Cell Metabolism
IF:20.565
发表时间:2016年
通信作者:Fredrik Backhed
通信作者单位:哥德堡大学分子和临床医学系和心血管代谢研究中心瓦伦堡实验室
综述内容
引言
肠道菌群对健康和疾病的作用越来越受到重视。根据最初的发现,高脂饮食无法诱导无菌(GF)小鼠肥胖,而且这种表型可以通过正常肠道微生物群的定植来逆转,说明肠道微生物群已经成为促进宿主代谢的环境因素。肥胖是通过胰岛素抵抗引起的,与2型糖尿病(T2DM)、心血管疾病和非酒精性脂肪性肝病(NAFLD)及其并发症肝硬化和癌症等相关。不管是对人类还是在对小鼠的研究中,肠道微生物群在肥胖和相关疾病中都发生了改变。然而,潜在的分子机制和微生物诱导的信号通路仍然是未知的。肠道微生物群具有产生生物活性化合物的很大潜力,其可通过激活各种细胞中的同源受体向宿主发出信号。一类重要的微生物产生的代谢物是胆汁酸,由肝脏中的胆固醇合成的内源性分子,其通过肠道微生物群进一步代谢后激活肠道、肝脏和外周的受体调节包括代谢过程的几个宿主过程。
人体产生的主要胆汁酸是鹅去氧胆酸(CDCA)和胆酸(CA),而啮齿动物产生CA和鼠胆酸(MCAs),主要是β-MCA(βMCA)。在排泄到胆汁并进一步通向十二指肠之前,胆汁酸与氨基酸甘氨酸酰胺化,并在较小程度上与人体中的牛磺酸酰胺化。相反,胆汁酸几乎完全与小鼠和大鼠中的牛磺酸结合。饭后胆汁酸释放到十二指肠中,然后在回肠中重新吸收结合的胆汁酸通过门静脉血再循环到肝脏,这个过程被称为肠肝循环并保留了超过95%的胆汁酸池。在最近发表的一篇优秀综述中介绍了胆汁酸化学和生理学的细节。从这个角度来看,我们总结了微生物群如何调节胆汁酸以及这种调节如何导致代谢疾病。
1 微生物调节胆汁酸的合成
胆汁酸的形成是复杂的,包括由至少17种不同酶催化的几个反应步骤。胆汁酸在肝脏中合成,可通过两种不同的途径完成(图1),经典(或中性)途径在正常条件下占胆汁酸产生的比例至少75%,由胆固醇7α-羟化酶 (CYP7A1)催化的胆固醇的7α-羟基化引发。CYP7A1是限速酶,决定了产生的胆汁酸的量。替代(或酸性)途径由甾醇-27-羟化酶(CYP27A1)引发,形成的27-羟基胆固醇通过氧固醇7α-羟化酶(CYP7B1)进一步羟基化。由胆固醇和氧固醇产生的7α-羟基化中间体经过甾醇环修饰和侧变氧化缩短反应步骤。我们之前的研究已经表明了肠道微生物群调节了几种酶的表达,包括CYP7A1、CYP7B1和CYP27A1。
值得注意的是,尽管替代途径主要产生CDCA,但经典途径产生CDCA和CA这两种原发性胆汁酸,它们的比例由合成CA所必需的酶-甾醇12α-羟化酶(CYP8B1)决定,而且我们已经证明了这种酶不受微生物调节。除CA和CDCA外,小鼠还产生MCAs和熊去氧胆酸(UDCA)作为原发性胆汁酸。而在人类中,UDCA是二级胆汁酸,通常不检测MCAs。我们和其他人已经证明,在没有细菌的情况下(如在GF或抗生素治疗的小鼠或大鼠中),胆汁酸池主要由初级结合的胆汁酸组成,GF大鼠的早期实验表明,MCCA在肝脏中由CDCA合成,然而,该过程中涉及的酶仍未定义。有趣的是,微生物群在肝脏中的MCAs合成中具有强调节作用,可能也调节未知合成酶。
肝脏在两步反应中将胆汁酸C-24位与甘氨酸或牛磺酸结合。牛磺酸以及胆汁酸酰基辅酶A合成所需的酶,其是也是胆汁酸共轭所需的两种酶中的第一种,这种酶也是由微生物调节的。共轭胆汁酸通过胆汁盐输出泵(BSEP)主动转运到胆汁中,并且可以将其储存在胆囊内,直到摄取膳食后释放到十二指肠。有趣的是,GF小鼠胆汁酸池中的tauro-βMCA(TβMCA)比例增加(~80%),而常规饲养(CONV-R)小鼠的比例为~50%。大约95%的胆管分泌的胆汁酸从肠道重新吸收,主要是作为结合胆汁酸在回肠末端通过钠依赖性胆汁酸转运蛋白(ASBT,也称为IBAT),并通过门静脉再循环至肝脏又被分泌出来,这个过程称为肠肝循环,每天大约发生六次。
早期的研究表明,GF大鼠的回肠粘膜具有增加的吸收tauro-CA(TCA)的能力,导致与CONV-R大鼠相比,GF的半衰期延长4至5倍。加上我们发现ASBT受微生物调节,这也证明了肠道微生物群不仅可以调节胆汁酸合成而且还可以调节胆汁酸摄取,这两者都有助于GF小鼠中形成较大的胆汁酸池。
2 胆汁酸的微生物代谢
微生物去缀合(即去除甘氨酸或牛磺酸缀合物)防止通过ASBT从小肠中主动再摄取。胆汁酸去缀合由具有胆汁盐水解酶(BSH)活性的细菌进行。宏基因组学分析表明功能性BSH存在于人类肠道中的所有主要细菌和古菌中,包括乳酸杆菌、双歧杆菌、梭菌和拟杆菌的成员。与其他微生物生态系统相比,BSH在肠道微生物中富集,而且与胆汁毒性的抗性增加有关。通过ASBT逃避摄取的非结合原代胆汁酸进入结肠,它们通过7-脱羟基作用代谢成二级胆汁酸(图1)、来自CDCA的石胆酸(LCA)和来自CA的脱氧胆酸(DCA)。对原代鼠αMCA和βMCA的相同反应导致鼠脑磷酸(MDCA)的形成。Omega-MCA(ωMCA)是βMCA的主要代谢产物,由6β-差向异构化形成。βMCA的其他代谢产物是通过6β-差向异构化和额外的7β-脱羟基作用形成的猪去氧胆酸(HDCA),以及通过6β-差向异构化和进一步的7β-差向异构化形成的猪胆酸(HCA;也称为γMCA)。在人类中,UDCA是由梭状芽胞杆菌将CDCA的7α/β-异构化形成。进一步重要的异构化分别是3α/β-羟基和5-H-β/α-异胆汁酸的异构化。
胆汁酸的微生物代谢增加了多样性以及胆酸池的疏水性,有利于胆汁酸的排泄,总共约5%。去缀合的次级胆汁酸的一小部分也通过被动扩散从肠道吸收,并在肠肝循环中富集,然后充当宿主中的信号分子。在小鼠粪便中检测到的ωMCA,MDCA和HDCA水平在不同的小鼠中可能有很大差异,这可能归因于肠道微生物群组成的差异改变了胆汁酸的微生物修饰。重要的是已经有人提出,人类肠道微生物群不能代谢βMCA,小鼠和人类的有不同的胆汁酸谱,可能影响胆汁酸受体的信号传导,因此在将小鼠的观察结果发现转化为人类时需要谨慎。
7-脱羟基的复杂过程包括由细菌与胆汁酸诱导的基因进行的许多反应。能够产生二级胆汁酸的细菌已经鉴定了两种属于厚壁杆菌的属分别为梭菌(簇XIVa和XI)和真细菌中。有趣的是, DCA水平的增加与小鼠的肥胖和癌症有关,进一步支持了微生物的胆汁酸转化是宿主代谢重要作用。
胆汁酸的另一种主要微生物转化是生成氧-(或酮-)胆汁酸,通过在环位置3,7或12的羟基氧化产生这些羟基是由细菌与羟类固醇脱氢酶(HSDHs)催化的,羟基类固醇脱氢酶(HSDHs)在放线菌、变形菌、厚壁杆菌和拟杆菌中都存在。这些氧化反应是可逆的并且最终可导致差向异构化。在常规和高级胆汁酸分析中,由于合适的参考化合物的可用性有限,所形成的化合物通常被忽略。值得注意的是,异胆汁酸存在于人类血清和尿液中,特别是在结肠和粪便中,其中iso-LCA和iso-DCA是继LCA和DCA之后第二丰富的胆汁酸,在某些个体中浓度分别达到290和390 mM。具有产生异胆酸的能力的细菌包括迟缓真杆菌和产气荚膜梭菌,iso-DCA不仅由迟缓真杆菌产生,而且产气荚膜梭菌也能产生,而且有人指出,iso-DCA有利于拟杆菌属的生长,暗示微生物HSDH作为肠道微生物组成和宿主代谢的潜在调节剂,可能为微生物群如何调节宿主内的信号通路开辟新的途径。

图1 胆汁酸的合成与代谢
3 通过FXR调节胆汁酸合成:微生物群的影响
胆汁酸的合成受到核受体FXR的负反馈抑制的严格调节,FXR是一种与启动子区域结合并启动多种靶基因表达的转录因子,在几种组织中表达。FXR表达最高的组织也是研究最广泛的是肝脏和回肠。FXR在肾脏表达相当高的水平,而在心脏、卵巢、胸腺、眼睛、脾脏和睾丸中水平较低。在肝脏中,胆汁酸活化的FXR诱导小异二聚体伙伴(SHP)的表达,与肝受体同源蛋白-1 (LRH-1)结合,从而抑制Cyp7a1基因的表达(图2)。除了肝脏中的局部作用外,FXR还被远端回肠中的胆汁酸激活,在那里它诱导FGF15(人类中的FGF19)的表达。FGF15 /19通过门静脉血到达肝脏,并与FGF受体4(FGFR4)/b-klotho异二聚体复合物结合,触发JNK1/2和ERK1/2信号级联反应,同时抑制Cyp7a1的表达(图2)。组织特异性Fxr缺陷小鼠的研究表明回肠FXR激活导致Cyp7a1介导的胆汁酸合成的抑制比肝FXR激活更强,而肝FXR能调节Cyp8b1进而形成CA。
FXR最有效的配体是CDCA,其次是CA,DCA和LCA。UDCA不激活FXR,它反而抑制FXR活化,这与计算机模拟FXR结合研究一致。最近,小鼠牛磺酸结合的原代胆汁酸TαMCA和TβMCA已被鉴定为天然存在的FXR拮抗剂。GF小鼠有TβMCA的积累,FXR信号减少,胆汁酸合成增加,TβMCA在没有细菌的情况下不能代谢。我们已经证明了肠道微生物群的存在主要在回肠中降低了TβMCA的水平,导致Fgf15通过FXR活化的表达增加。因此,我们证明了微生物群以FXR依赖性方式调节胆汁酸的代谢和合成。相反,我们没有观察到Cyp8b1和CA生物合成的微生物调控。
Gonzalez小组证实了我们关于微生物群调节FXR信号传导的研究结果,他们使用抗氧化剂四甲基哌啶(4-羟基-2,2,6,6-四甲基哌啶1-氧基)来改变微生物群和胆汁酸谱,从而导致TβMCA水平升高和FXR信号受抑制。 具体来说,他们发现四甲基哌啶能降低乳杆菌属和梭菌属丰度(簇IV和XIV),这伴随着BSH活性降低和肠TβMCA的积累。最近,他们发现了βMCA的甘氨酸缀合物(GβMCA)也是FXR拮抗剂,与TβMCA相反,它对粪便微生物BSH具有抗性。 然而,目前还不知道TβMCA或GβMCA是否可以拮抗人类中的FXR信号传导,以及更适合甘氨酸缀合的胆汁酸的人类微生物群如何代谢GβMCA。
4 通过FXR微生物调节宿主代谢过程
胆汁酸在肝脏和小肠等代谢活性组织中作为信号分子和FXR表达的重要性,引起了人们对剖析FXR在代谢疾病中的作用的极大兴趣。然而,用缺乏fxr的小鼠进行的实验产生了部分相互矛盾的结果,显然饮食、以及动物设施之间微生物群的差异,可能会导致这些不同的表型。饮食中fxr缺乏的小鼠容易发生高血糖和高胆固醇血症,而在Ldlr-/-背景下喂食高脂饮食的fxr缺陷小鼠,改善了血脂,并对饮食诱导的肥胖和动脉粥样硬化的发展具有保护作用。相比之下,Apoe-/-基因背景下的fxr缺陷小鼠动脉粥样硬化增加。然而,总体而言,与对照组小鼠相比,fxr缺陷小鼠喂食高脂饮食或有遗传肥胖背景 (ob/ob)的小鼠似乎可以预防肥胖,并改善葡萄糖稳态。然而,FXR与特定营养素之间的相互作用值得进一步研究。
小鼠和人类的肠道微生态受到饮食的显著影响。微生物群不仅能代谢胆汁酸,还通过FXR影响信号传导。微生物群将结合FXR的拮抗剂TβMCA去偶联,从而促进小鼠中的FXR信号传导,并且还产生作为TGR5配体的二级胆汁酸。因此,使用GF和抗生素处理的小鼠的微生物群通过FXR信号传导诱导饮食诱导的肥胖。微生物群诱导脂肪组织炎症并增加基因在肝脏表达以FXR依赖的方式参与脂质摄取。除了信号传导途径的微生物调节,我们还注意到,用高脂肪喂养FXR缺陷型小鼠中的改变的肠道微生物移植到GF小鼠时,相对于正常组,体重增加减少,这表明FXR依赖性微生物信号传导可通过多种途径导致代谢受损。目前尚不清楚人类的FXR调节是否会导致微生物群的改变。
利用FXR的肠和肝损坏的研究表明,肝脏和肠都是FXR信号依赖性葡萄糖和脂质控制的中枢器官。Schmitt观察到FXR在肝脏中的表达可以防止肝脏脂肪变性以及甘油三酯和胆汁酸水平升高。相比之下,肠特异性FXR缺失的小鼠即使在喂食高脂饮食时,也不会发生肝脏脂肪变性和肥胖。因此,肠道和肝脏FXR可能对脂肪变性有相反的作用。因此,有一种观点认为FXR可能对正常饮食有益处,而在高脂肪饮食中,FXR缺乏可能会预防代谢疾病,特别是当肠道FXR受到抑制时。阐明肠道微生物群是如何影响人类小肠和肝脏中的FXR信号是非常重要的。
5 胆汁酸和FXR以饮食依赖的方式调节微生物群
微生物群和胆汁酸之间的相互作用并不是单向的。胆汁酸也可以通过促进胆汁酸代谢细菌的生长和抑制其他胆汁敏感细菌的生长来塑造肠道微生物群落。研究表明,阻止胆汁流入肠道的胆道阻塞导致细菌过度生长和细菌在小肠内易位,这种表型可以通过胆汁酸给药逆转。 这也证明了胆汁酸不仅能通过破坏细菌细胞膜发挥直接的抗菌作用,而且还间接通过FXR诱导抗微生物剂(如iNOS和IL-18)的转录,通过免疫系统影响肠道微生物群。
我们最近发现FXR可以调节微生物群,而且微生物群的变化与胆汁酸组成的变化有关。高脂饮食的fxr缺陷小鼠与相同饮食的WT小鼠相比,厚壁菌门的相对丰度增加,拟杆菌门的丰度减少。血清和盲肠中胆汁酸的谱图也随着TβMCA和βMCA水平的升高而改变,这可能是由于这些初级胆汁酸转化为次级胆汁酸的程度降低所致。fxr-缺陷小鼠体内原代胆汁酸的积累表明,在没有FXR的情况下,该微生物群可能降低了胆汁酸的代谢能力。GF fxr-缺陷小鼠的胆汁酸分布与WT相似,表明fxr对胆汁酸组成的影响依赖于微生物群的改变。
Devkota等人进一步证明了饮食、胆汁酸和微生物之间的强相互作用。用由牛奶脂肪或多不饱和脂肪组成的高脂食物喂养的老鼠,无论脂肪来源如何,高脂肪饮食增加了拟杆菌门的丰度,减少了厚壁菌门的丰度。胆汁酸和微生物之间存在着动态的相互作用,可以通过饮食来改变,从而对宿主代谢产生有利或有害的影响。
了解微生物群、饮食和胆汁酸之间的相互作用对于阐明微生物群如何调节代谢至关重要。Koeth等研究发现,添加肉碱或TMAO的饮食可以减少小鼠胆汁酸池,从而影响FXR信号,改变代谢表型。因此,微生物群、胆汁酸、FXR以及微生物和宿主代谢的共同代谢产物之间的相互作用可能导致代谢性疾病。
6 TGR5在宿主代谢中的作用
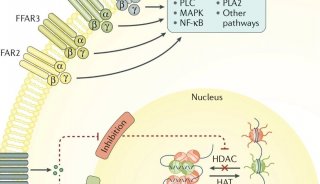
TGR5是另一种参与宿主代谢的胆汁酸反应受体。TGR5是一种结合性的G蛋白偶联受体,在胆囊、胎盘、肺、脾、肠、肝、棕白色脂肪组织、骨骼肌、骨髓中普遍高表达。TGR5主要由次级胆汁酸LCA和DCA激活,因此,在微生物与胆汁酸相互作用的背景下,它是一个有趣的靶点(图2)。
tgr5-缺陷小鼠是在2006年产生的,它们看起来健康而且具备生育能力,没有明显的异常。有趣的是,他们的胆汁酸池减少了,表明TGR5在胆汁酸稳态中的作用,但尚不清楚这是TGR5的直接影响还是通过FXR介导的。此外,tgr5缺乏的小鼠在喂食致石食物时,可以防止胆固醇胆结石的形成。
Watanabe等人发现,TGR5可能通过促进细胞内甲状腺激素活性从而增加棕色脂肪组织的产热,从而在能量平衡中发挥作用。他们发现高脂饮食诱导的肥胖和棕色脂肪组织中的脂肪堆积能够被膳食钙补充剂完全逆转,这表明FXR的依赖机制,但这被高活性合成FXR配体GW4064排除。也有大量的研究证实,当提供FXR的激动剂天然胆汁酸时,肠道微生物代谢可能产生TGR5的配体,强调了研究肠道微生物群的重要性。

图2 微生物对胆汁酸的修饰通过胆汁酸受体FXR和TGR5影响宿主代谢
7 NAFLD中微生物-胆汁酸相互作用的调节
NAFLD是代谢综合征的肝脏表现,是脂质和糖代谢失调的结果。NAFLD是一种复杂的疾病,从单纯性脂肪变性到非酒精性脂肪性肝炎(NASH),伴有纤维化、肝硬化,最后是肝癌。积累的数据表明,微生物群在NAFLD的发生发展中起作用。NAFLD患者FXR表达降低,血清甘油三酯水平升高。
目前对于NAFLD/NASH患者除了改善饮食和生活方式外,没有其他治疗方法,但是胆汁酸受体激活是目前争论最激烈的新治疗方法之一,包括半合成胆汁酸衍生物贝替胆酸(OCA)。与CDCA相比,OCA对FXR的配体结合力是CDCA的100倍,并且已经在NASH患者、有T2DM和无T2DM患者的2期和3期试验中进行了测试。与安慰剂组相比,NAFLD和T2DM患者使用OCA治疗6周,剂量依赖性胰岛素敏感性改善,体重减轻。一项多中心、双盲、随机、安慰剂对照、3a期研究表明,在有和没有T2DM的283名NASH患者中,OCA (FLINT)联合使用,72周的治疗可以显著改善NAS (NASH活动得分)。与对照组相比,OCA治疗组纤维化程度也有所改善。我们正在进行的由研究者发起的与OCA的试验旨在解释FXR在NAFLD患者体内激活的分子作用以及OCA对肠道菌群的影响。
8 减重手术后微生物-胆汁酸相互作用的变化
减肥手术是目前治疗肥胖症最有效的方法。最近我们证明了减肥手术对肠道菌群组成和功能有长期影响。与未手术的肥胖对照患者相比,减肥手术患者体内的微生物群发生了改变,移植到GF小鼠后从而减少了GF小鼠体内脂肪的增加。此外,与之前的研究一致的是,Rouxen-Y胃旁路术(RYGB)患者与肥胖对照组相比,餐后胆汁酸和FGF19反应增加。我们还发现FXR对小鼠减肥手术后的血糖代谢有重要贡献。相比之下,对TGR5缺陷小鼠的实验表明,在垂直袖状胃切除术(VSG)后,TGR5并没有导致体重减轻、能量消耗增加或葡萄糖刺激胰岛素分泌增加。然而,在TGR5缺陷小鼠中,糖耐量、肝胰岛素信号转导以及VSG后胆汁酸谱的代谢有利的变化均被减弱,表明TGR5至少对减肥手术后的一些有益作用有影响。
Flynn等人最近的一项研究显示,在胆汁转移(通过将胆汁酸重新释放到回肠)后,小鼠在体重减轻和葡萄糖耐受方面与RYGB手术后的小鼠相似。胆汁转移后,血清TβMCA和TωMCA水平显著升高,FXR信号量也显著降低,这与我们之前在小鼠VSG后诱导FXR的发现相反。这强调了FXR作为宿主代谢调节剂的作用的复杂性,其中通过减少和以特定于环境的方式增加FXR信号传导可以实现有益效果。 此外,在人体中减肥手术后肠道FXR信号传导如何被调节还有待证明。
9 结论
胆汁酸受体FXR和TGR5已成为代谢疾病的转化和介入研究的主要靶标。很明显,微生物群可通过胆汁酸的修饰调节FXR和TGR5的信号传导。因此,了解肠道微生物群及其对胆汁酸代谢和信号传导的影响可以提供对临床前数据解释的理解。然而,人和鼠胆汁酸具有显着不同的信号传导性质,并且缺乏丰富和/或表征的胆汁酸代谢物可能另外妨碍正确的结论。微生物群的差异也可以解释个体对胆汁酸衍生药物的反应,对微生物区系的更深入的了解可以识别出对治疗有积极反应的个体和有不良反应的个体。因此,针对微生物群,胆汁酸和FXR和/或TGR5信号之间的相互作用似乎发展成为治疗代谢疾病的有希望的途径,但需要更多的研究。
研究评论
本文总结了胆汁酸与肠道菌群之间的相互作用,阐述了FXR以及TGR5作为胆汁酸受体对代谢性疾病研究的重要性,首先胆汁酸可以影响肠道菌群,反过来肠道菌群又可以通过胆汁酸的修饰调节FXR和TGR5的信号传导,这对代谢性疾病的临床前数据解释至关重要。然而,作者也一直强调的是,目前大多数研究都是以小鼠为实验对象,而人和鼠胆汁酸具有显着不同的信号传导性质,不可全然照搬。因此,以FXR、TGR5为靶点,研究胆汁酸、微生物群之间的关系从而研究代谢性疾病,是一个很有希望的突破口,但仍然需要更多的研究。评论仅属小编一己之见,希望大家多提意见,提携小编进步。



长
按
关
注
解锁更多更精彩的「代谢组学」相关资讯!
微信公众号:麦特绘谱
Tel:400-867-2686
Web: www.metaboprofile.com