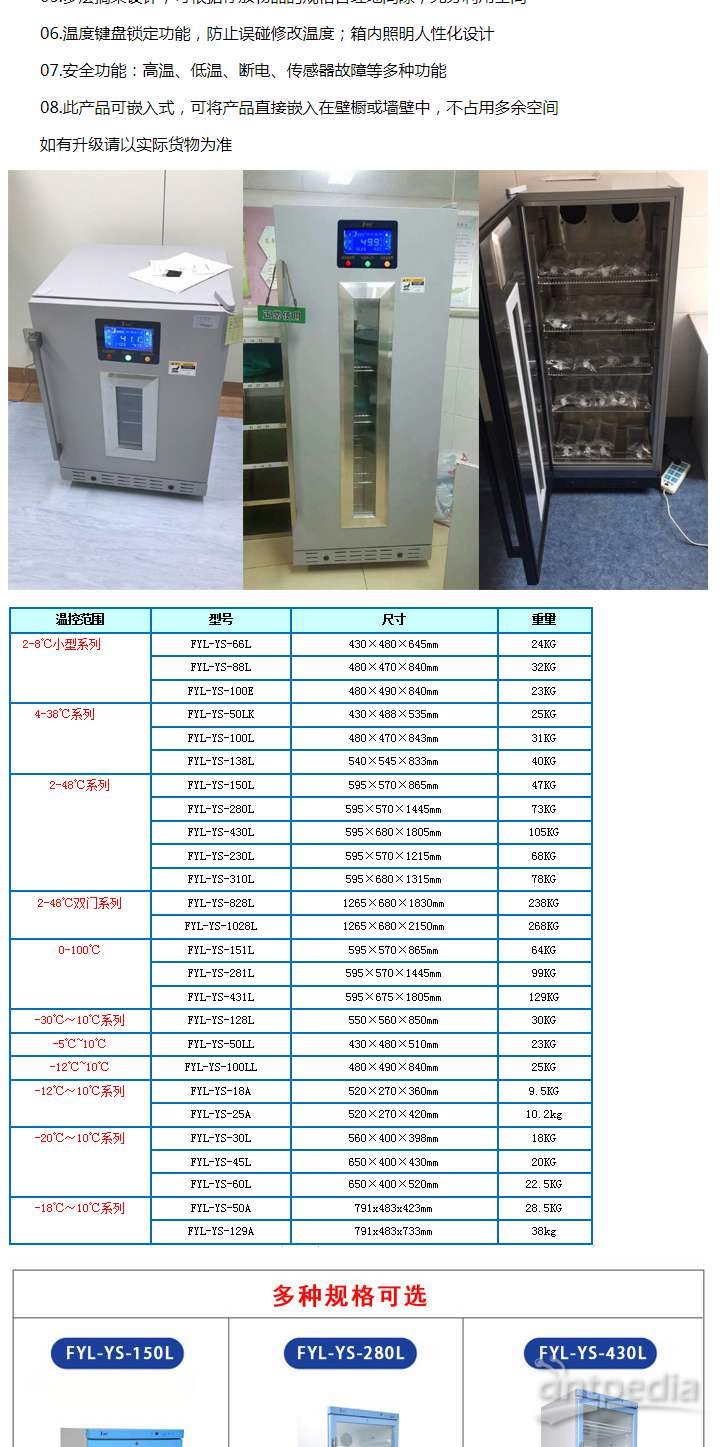
人源抗体开发——B细胞永生化 (二)
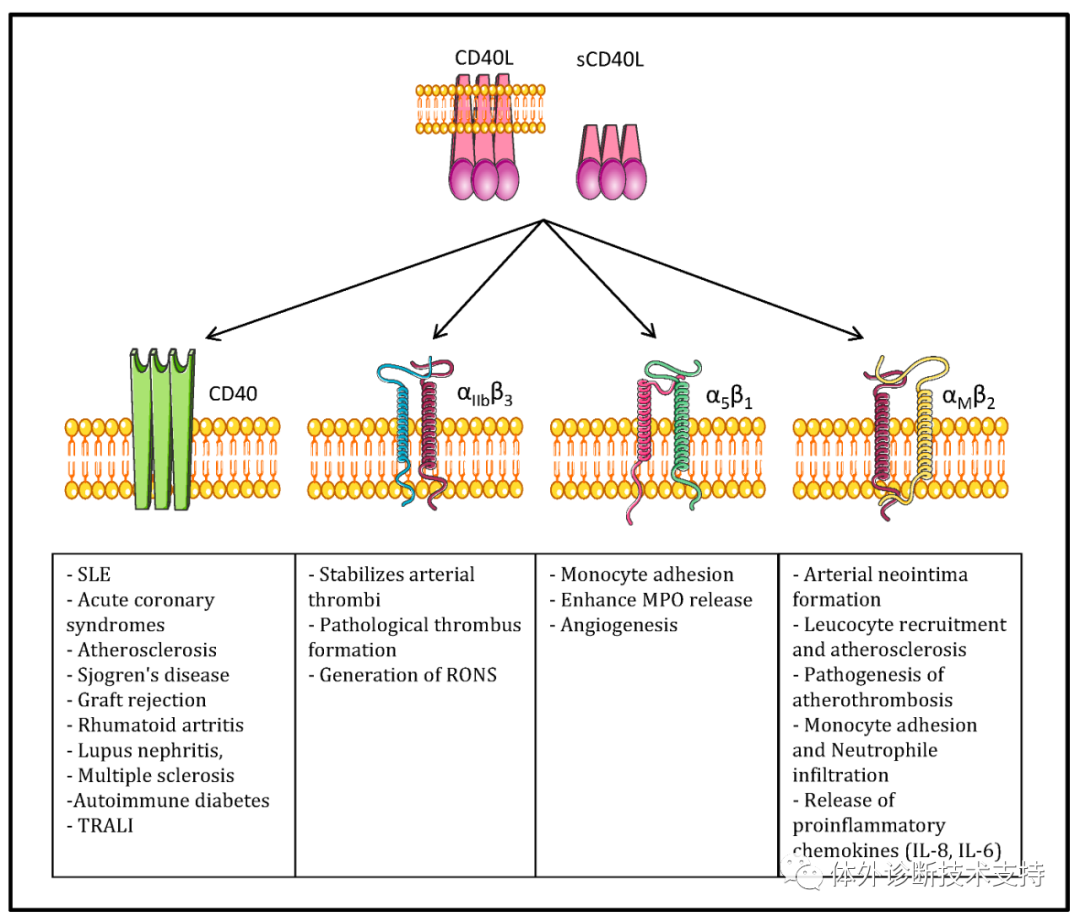

CD40 可通过活化NF-κB/ Rel 来诱导胚系R1 的转录, 起到IgG 同种异型的转换。 CD40L-CD40 结合后, 可诱导内源性TGF-β 及IL-10 表达, 促成VHDJH-Ca1 和VHDJH-Ca2 的转录及IgA 的分泌, 在IL-4 的存在下, CD40L 可诱导IgE 转录及表达。 通过CD40传导信号活化的NF-κB 可激活磷酸络氨酸激酶(PTK)和丝氨酸/素氨酸激酶, 这在CD40介导的B细胞活化中很关键.
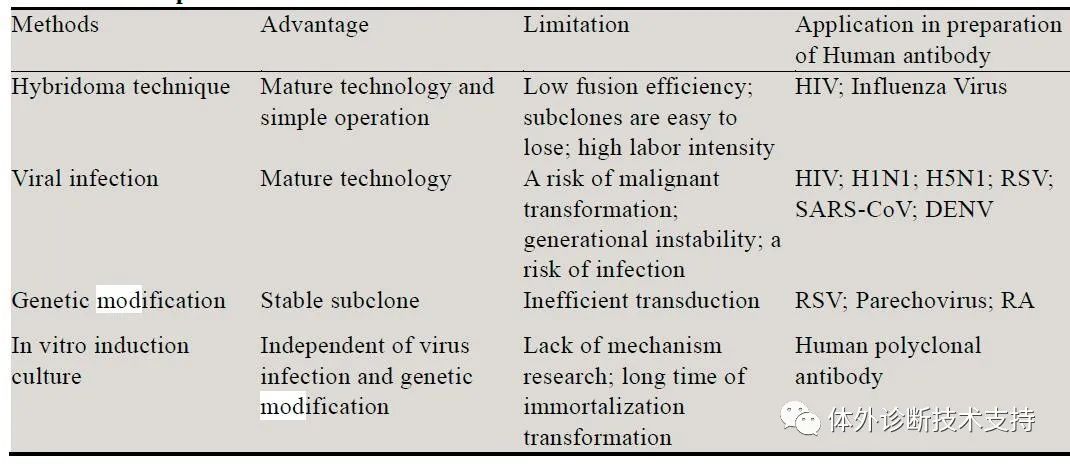
推荐

关注公众号
手机查看