DNA与蛋白质相互作用的研究方法有哪些
研究蛋白质与DNA相互作用的主要方法
一、引言
在许多的细胞生命活动中,例如DNA复制、mRNA转录与修饰以及病毒的感染等都涉及到DNA与蛋白质之间的相互作用的问题。
重组DNA技术的发展,人们已分离到了许多重要的基因。现在的关键问题是需要揭示环境因子及发育信号究竟是如何控制基因的转录活性。为此需要:
a、鉴定分析参与基因表达调控的DNA元件;
b、分离并鉴定这些顺式元件特异性结合的蛋白质因子;
这些问题的研究都涉及到DNA与蛋白质之间的相互作用。
研究DNA-蛋白质相互作用的实验方法主要包括:
a、凝胶阻滞实验; b、DNase 1 足迹实验;
c、甲基化干扰实验; d、体内足迹实验; f、拉下实验。
二、凝胶阻滞实验
1、概念:
凝胶阻滞实验(Gel retardation assay),要叫做DNA迁移率变动试验(DNA mobility shift assay)或条带阻滞实验(Band retardation assay)是在八十年代初期出现的用于在体外研究DNA与蛋白质相互作用的一种特殊的凝胶电泳技术。
2、原理:
在凝胶电泳中,由于电场的作用,裸露的DNA分子向正电极移动距离的大小是同其分子量的对数成反比。如果某种DNA分子结合上一种特殊的蛋白质,那么由于分子量的加大它在凝胶中的迁移作用便会受到阻滞,于是朝正极移动的距离也就相应的缩短,因而在凝胶中出现滞后的条带,这就是凝胶阻滞实验的基本原理。
3、过程:
1) 首先制备细胞蛋白质提取物(理论上其中含有某种特殊的转录因子)
2) 用放射性同位素标记待检测的DNA片段(含有转录因子的结合位点)
3) 这种被标记的探针DNA同细胞蛋白质提取物一起进行温育,于是产生DNA-蛋白质复合物
4) 在控制使DNA-蛋白质保持结合状态的条件下,进行非变性聚丙烯酰胺凝胶电泳
5) 最后进行放射自显影,分析电泳结果
4、实验结果的分析:
a、如果有放射性标记的条带都集中于凝胶的底部,这就表明在细胞提取物中不存在可以同探针DNA相互结合的转录因子蛋白质;
b、如果在凝胶的顶部出现放射性标记的条带,这就表明细胞提取物存在可与探针DNA结合的转录因子蛋白质。
5、DNA竞争实验:
DNA竞争实验(DNA competitive assay)的具体做法如下:
在DNA-蛋白质结合的反应体系中加入了超量的非标记的竞争DNA(competitor DNA),如果它同探针DNA结合的是同一种转录因子蛋白质,那么由于竞争DNA与探针DNA相比是极大超量的,这样绝大部分转录因子蛋白质都会被竞争结合掉,而使探针DNA仍然处于自由的非结合状态,可以在电泳凝胶的放射自显影图片上就不会出现阻滞的条带;
如果反应体系中加入的竞争DNA并不能同探针DNA竞争结合同一种转录因子,结果在电泳凝胶中的放射自显影图片上就会出现阻滞的条带。
6、应用:
a、凝胶阻滞实验可以用于鉴定在特殊类型细胞蛋白质提取物中,是否存在能同某一特定的DNA(含有转录因子结合位点)结合的转录因子蛋白质;
b、DNA竞争实验可以用来检测转录因子蛋白质同DNA结合的精确序列部位;
c、通过竞争DNA中转录因子结合位点的碱基突变可以研究此种突变竞争性能及其转录因子结合作用的影响;
d、也可以利用DNA同特定转录因子的结合作用通过亲和层析来分离特定的转录因子。
三、足迹实验
1、定义:
足迹实验(foot-printing assay),是一种用来检测被特定转录因子蛋白质特异性结合的DNA序列的位置及其核苷酸序列结构的专门实验方法。
2、原理:
当DNA分子中的某一区段同特异的转录因子结合之后便可以得到保护而免受DNaseI 酶的切割作用,而不会产生出相应的切割分子,结果在凝胶电泳放射性自显影图片上便出现了一个空白区,俗称为“足迹”。
3过程:
将待检测的双链DNA分子在体外用32P作5‘末端标记,并用适当的限制性内切酶切出其中的一个末端,于是便得到了一条单链末端标记的双链DNA
在体外同细胞蛋白质提取物(细胞核提取物也可以)混合,形成DNA-蛋白质复合体
在反应混合物中加入少量的DNase I,并控制用量使之达到平均每条DNA链,只发生一次磷酸二酯键的断裂:
a、如果蛋白质提取物中不存在与DNA结合的特定蛋白质,使DNase I消化之后,便会产生出距离放射性标记末端1个核苷酸,2个核苷酸,3个核苷酸------等等一系列前后长度均相差一个核苷酸的不间断的连续的DNA片段梯度群体;
b、如果DNA分子同蛋白质提取物中的某种转录因子结合,被结合部位的DNA就可以得到保护免受DNase I酶的降解作用;
除去蛋白,加样在20%序列胶上进行电泳分离,实验分两组:
a、实验组:DNA+蛋白质混合物
b、对照组:只有DNA,未与蛋白质提取物进行温育
最后进行放射性自显影,分析实验结果。
4、结果判断:
实验组凝胶电泳显示的序列,出现空白的区域表明是转录因子蛋白质结合部;与对照组序列比较,便可以得出蛋白质结合部位的DNA区段相应的核苷酸序列。
5、其他的足迹实验方法:
除了DNase1足迹试验之外,目前还发展出了若干种其他类型的足迹实验,例如:
a、 自由羟基足迹实验;b、菲咯啉铜足迹实验;c、DMS(硫酸二甲酯)足迹实验
DMS(硫酸二甲酯)足迹实验的原理
DMS能够使DNA分子中裸露的鸟嘌呤(G)残基甲基化,而六氢吡啶又会对甲基化的G残基作特异性的化学切割。如果DNA分子中某一区段同转录因子结合,就可以避免发生G残基的甲基化而免受六氢吡啶的切割作用。
四、甲基化干扰实验
1、概念:
甲基化干扰实验(Methylation interference assay)是根据DMS(硫酸二甲酯)能够使DNA分子中裸露的鸟嘌呤(G)残基甲基化,而六氢吡啶又会对甲基化的G残基作特异性的化学切割这一原理设计的另一种研究蛋白质同DNA相互作用的实验方法。
应用这种技术可以检测靶DNA中G残基的优先甲基化,对尔后的蛋白质结合作用究竟会有什么效应,从而更加详细的揭示出DNA与蛋白质相互作用的模式。
2、实验步骤:
用DMS处理靶DNA使之局部甲基化(平均每条DNA只发生一个G碱基甲基化作用)
同细胞蛋白质提取物一起进行温育,促进使DNA与蛋白质的结合
进行凝胶电泳形成两种靶DNA条带:
a、 其一没有同蛋白质结合的DNA正常电泳条带
b、其二同特异蛋白质结合而呈现滞后的DNA电泳条带
将这两种DNA电泳条带分别从凝胶中切出,并用六氢吡啶进行切割,结果为:
a、甲基化的G残基被切割:因为转录因子蛋白质只能够同未发生甲基化的正常的结合位点结合,所以在转录因子DNA结合位点序列中的G残基如果被DMS甲基化之后,转录因子就无法同其结合位点(顺式元件)发生结合作用,从而使得结合位点中的G残基同样也要被六氢吡啶切割;
b、不具有甲基化G残基的靶DNA 序列则不会被切割
将结合蛋白质的DNA条带和不结合蛋白质的DNA条带,经六氢吡啶切割作用之后,再进行凝胶电泳
作放射自显影,读片并分析结果
3、结果判断:
a、同转录因子蛋白质结合的靶DNA序列,经六氢吡啶切割之后,电泳分离呈现两条带,有一个空白区
b、不同转录因子蛋白质结合的靶DNA序列,经六氢吡啶切割后,电泳分离呈现三条带,没有空白区域的出现。
4、应用:
a、甲基化干扰实验可以用来研究转录因子与DNA结合位点中的G残基之间的联系;
b、是足迹实验的一种有效的补充手段,可以鉴定足迹实验中DNA与蛋白质相互作用的精确位置
5、缺点:
DMS只能使DNA序列中的G和A残基甲基化,而不能使T和C残基甲基化。
五、体内足迹实验
上面讨论的三种研究转录因子与DNA相互作用的方法,有一个共同的不足之处在于它们是在体外进行的实验,因此人们就会考虑这些实验结果是否能够反映细胞内发生的真实生命过程,即细胞内发生的真实的DNA与蛋白质的相互作用情况。
为了解答这个问题,科学家就设计出了一种体内足迹试验(in vivo foot-printing assay),该方法可以看做是体外DMS足迹实验的一个变种。
1、原理:
体内足迹试验的原理原则上同体外DMS足迹实验无本质差别,即
a、DMS能够使G残基甲基化;
b、六氢吡啶能特异的切割甲基化的G残基;
c、同特异转录因子蛋白质结合的识别序列中的G残基由于受到蛋白质的保护而不会被DMS甲基化,于是不会被六氢吡啶切割;
d、同对照的裸露的DNA形成的序列梯作比较,就会发现活细胞DNA形成的序列梯中缺少G残基没有被切割的相应条带。
2、过程:
用有限数量的化学试剂DMS处理完整的游离细胞,使渗透到胞内的DMS浓度恰好导致天然染色体DNA的G残基发生甲基化
对这些经过DMS处理的细胞提取DNA,并在体外加入六氢吡啶作消化反应
PCR扩增后作凝胶电泳分析,因为在体外实验中用的是克隆的DNA片段其数量足够,而在体内足迹实验中用的是从染色体DNA中分离获得的任何一种特异的DNA,其数量是微不足道的,所以需要经PCR扩增以获得足够数量的特异DNA
放射自显影,读片并记录读片的结果
3、结果判断:
a、能够同转录因子蛋白质结合的DNA区段其中G残基受到保护因而不会被DMS甲基化避免了六氢吡啶的切割作用;
b、体外裸露的DNA分子上,G残基被DMS甲基化而被六氢吡啶切割。
六、拉下实验(Pull-down assay)
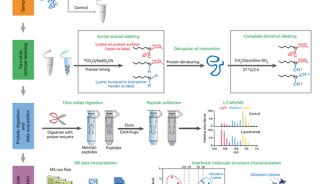
拉下实验又叫做蛋白质体外结合实验(binding assay in vitro),是一种在试管中检测蛋白质之间相互作用的方法。其基本原理是将某种小肽(例如生物素、6-His标签以及谷胱甘肽转移酶等)的编码基因与诱饵蛋白的编码基因重组,表达为融合蛋白。分离纯化融合蛋白并与磁珠结合,使之固相化之后,再与表达目的蛋白的细胞提取物混合保温适当时间,例如在4℃下保温过夜,使目标蛋白同已经固定在磁珠表面的融合蛋白中的诱饵蛋白充分的结合。离心收集与固定化的融合蛋白(即与磁珠相互结合的融合蛋白)中的诱饵蛋白相结合的目的蛋白,经过煮沸处理使目的蛋白与诱饵蛋白相脱离从而从固相支持物(例如磁珠)上脱离下来,收集样品,再与目标蛋白的抗体作Western blotting分析,以检测出与诱饵蛋白的目标的目标蛋白。
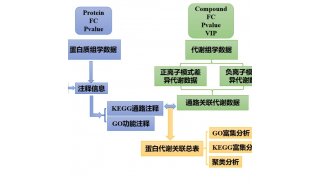
染色质免疫共沉淀技术(ChIP) 真核生物的基因组DNA以染色质的形式存在。因此,研究蛋白质与DNA在染色质环境下的相互作用是阐明真核生物基因表达机制的基本途径。染色质免疫沉淀技术(chromatin immunoprecipitation assay, CHIP)是目前唯一研究体内DNA与蛋白质相互作用的方法。它的基本原理是在活细胞状态下固定蛋白质-DNA复合物,并将其随机切断为一定长度范围内的染色质小片段,然后通过免疫学方法沉淀此复合体,特异性地富集目的蛋白结合的DNA片段,通过对目的片断的纯化与检测,从而获得蛋白质与DNA相互作用的信息。CHIP不仅可以检测体内反式因子与DNA的动态作用,还可以用来研究组蛋白的各种共价修饰与基因表达的关系。而且,CHIP与其他方法的结合,扩大了其应用范围:CHIP与基因芯片相结合建立的CHIP-on-chip方法已广泛用于特定反式因子靶基因的高通量筛选;CHIP与体内足迹法相结合,用于寻找反式因子的体内结合位点;RNA-CHIP用于研究RNA在基因表达调控中的作用。由此可见,随着CHIP的进一步完善,它必将会在基因表达调控研究中发挥越来越重要的作用。 染色体免疫共沉淀(Chromatin Immunoprecipitation,ChIP)是基于体内分析发展起来的方法,也称结合位点分析法,在过去十年已经成为表观遗传信息研究的主要方法。这项技术帮助研究者判断在细胞核中基因组的某一特定位置会出现何种组蛋白修饰。ChIP不仅可以检测体内反式因子与DNA的动态作用,还可以用来研究组蛋白的各种共价修饰与基因表达的关系。近年来,这种技术得到不断的发展和完善。采用结合微阵列技术在染色体基因表达调控区域检查染色体活性,是深入分析癌症、心血管疾病以及中央神经系统紊乱等疾病的主要代谢通路的一种非常有效的工具。 它的原理是在保持组蛋白和DNA联合的同时,通过运用对应于一个特定组蛋白标记的生物抗体,染色质被切成很小的片断,并沉淀下来。IP是利用抗原蛋白质和抗体的特异性结合以及细菌蛋白质的“prorein A”特异性地结合到免疫球蛋白的FC片段的现象活用开发出来的方法。目前多用精制的prorein A预先结合固化在argarose的beads上,使之与含有抗原的溶液及抗体反应后,beads上的prorein A就能吸附抗原达到精制的目的。实验最需要注意点就是抗体的性质。抗体不同和抗原结合能力也不同,免染能结合未必能用在IP反应。建议仔细检查抗体的说明书。特别是多抗的特异性是问题。其次,要注意溶解抗原的缓冲液的性质。多数的抗原是细胞构成的蛋白,特别是骨架蛋白,缓冲液必须要使其溶解。为此,必须使用含有强界面活性剂的缓冲液,尽管它有可能影响一部分抗原抗体的结合。另一面,如用弱界面活性剂溶解细胞,就不能充分溶解细胞蛋白。即便溶解也产生与其它的蛋白结合的结果,抗原决定族被封闭,影响与抗体的结合,即使IP成功,也是很多蛋白与抗体共沉的悲惨结果。再次,为防止蛋白的分解,修饰,溶解抗原的缓冲液必须加蛋白每抑制剂,低温下进行实验。每次实验之前,首先考虑抗体/缓冲液的比例。抗体过少就不能检出抗原,过多则就不能沉降在beads上,残存在上清。缓冲剂太少则不能溶解抗原,过多则抗原被稀释。 ChIP的一般流程: 甲醛处理细胞---收集细胞,超声破碎---加入目的蛋白的抗体,与靶蛋白-DNA复合物相互结合---加入ProteinA,结合抗体-靶蛋白-DNA复合物,并沉淀---对沉淀下来的复合物进行清洗,除去一些非特异性结合---洗脱,得到富集的靶蛋白-DNA复合物---解交联,纯化富集的DNA-片断---PCR分析。 在PCR分析这一块,比较传统的做法是半定量-PCR。但是现在随着荧光定量PCR的普及,大家也越来越倾向于Q-PCR了。此外还有一些由ChIP衍生出来的方法。例如RIP(其实就是用ChIP的方法研究细胞内蛋白与RNA的相互结合,具体方法和ChIP差不多,只是实验过程中要注意防止RNase,最后分析的时候需要先将RNA逆转录成为cDNA);还有ChIP-chip(其实就是ChIP富集得到的DNA-片段,拿去做芯片分析,做法在ChIP的基础上有所改变,不同的公司有不同的做法,要根据公司的要求来准备样品)。
GST沉淀实验(GST-pull down实验)(细胞外蛋白质相互作用)5-11
上面提到,用酵母双杂交方法筛选到的蛋白需要作进一步的鉴定。鉴定方法之一就是 GST
沉淀实验。GST 沉淀实验主要是用来证明蛋白质胞外的相互作用。蛋白质在胞外的相互作用排除了酵母细胞内复杂体系的干扰,,比较直接地检验蛋白质分子之间存在的物理的相互作用。同酵母双杂交实验一样,运用此法也可以证明相互作用的蛋白分子中是否有参与调节作用的结构域或 motif。GST 沉淀实验原理就是,把你要研究的蛋白基因亚克隆到带有GST(谷胱甘肽转移酶)基因的原核表达载体中,并在细菌中表达 GST 融合蛋白(GST-X)。把 GST 融合蛋白挂到带有 GST 地物的 Sepharose beads 上,然后把另一种蛋(Y)白加入其中。由于蛋白质之间的结合作用,形成了这样的复合物:GST-X----Y。这一复合物与固体支持物(Sepharose beads)又
结合在一起,可以被沉淀下来。此法又有在不同情况下的具体应用,以下一一作介绍。
(1) GST 融合蛋白与重组蛋白的相互作用
GST 融合蛋白是在原核表达的,所以没有经过过多的象真核细胞内具有的蛋白修饰作用。
所以另一种用来检验相互作用的蛋白也可以用原核表达出来,也就是所谓的重组蛋白。当 GST融合蛋白把重组蛋白沉淀下来,然后用重组蛋白的抗体作 Western blotting 检测。
(2) GST 融合蛋白与体外 TNT 系统合成的多肽或蛋白的相互作用
用来检验与GST融合蛋白相互作用的蛋白或多肽也可以用TNT体外蛋白合成体系进行合
成,并且还可以在要合成的蛋白或多肽N端或C端加上便于检测的标签。GST融合蛋白沉淀下来的蛋白或多肽可以用该蛋白或多肽的抗体或标签抗体进行Western blotting检测。如果蛋白之间结合力非常弱,用Western blotting检测方法难以检测到,你可以在TNT体外合成时给蛋白进行同位素(S32)标记。这样沉淀下来的蛋白进行放射自显影,检测灵敏度将极大提高。
(3) GST 融合蛋白与细胞内源性蛋白质的相互作用
GST融合蛋白还可以把细胞提取物中有相互作用的内源性蛋白质沉淀下来。如果内源性蛋
白含量低或结合力弱,可以采用脉冲法使细胞在某一段时间内合成的所有蛋白质都标记上放射性同位素(S32),然后提取细胞总蛋白与GST融合蛋白温育。GST融合蛋白沉淀下的带有放射性标记的蛋白跑电泳,进行放射自显影。
(4) GST 融合蛋白与细胞内瞬时表达的蛋白质的相互作用
当内源性蛋白质含量低,并且有可能影响蛋白质的相互作用,也可以把该蛋白的基因转染
到靶细胞内进行过表达,然后检验蛋白质相互作用。
(5) GST 融合蛋白与待测蛋白的相互作用有可能与待测蛋白的磷酸化状态有关在进行 GST 沉淀实验时,有时也会遇到比较复杂的情况,具体情况具体分析,分别对待。比如,两个蛋白质之间发生相互作用时有可能与蛋白磷酸化状态有关。或者蛋白首先被磷酸化后方能产生相互作用,或者磷酸化的蛋白必需脱磷酸化后才能产生相互作用。如果你确实在你
的实验中发现了其中一种情况,这将是一个非常有意义的结果。
3, 免疫共沉淀(co-immunoprecipitation )(细胞内蛋白质相互作用)9-16
免疫共沉淀技术用来证明蛋白质在胞内是否有相互作用。一般来说,两种蛋白在细胞内发
生相互作用时会形成两种蛋白的复合物,这样就可以先用一种蛋白的抗体把免疫复合物沉淀下来,然后用另一种蛋白的抗体进行 Western blotting 检测,看两种蛋白之间是否确实形成免疫复合物,并能与 protein A/G agarose 一起沉淀下来。免疫共沉淀原理简单,但技术极为复杂。因为细胞内蛋白种类繁多,制约因素多。如果两种蛋白之间可以发生相互作用,并不是这两种蛋白所有分子都参与结合作用,也可能只有极少部分蛋白分子结合在一起(足以满足细胞功能需要)。在提取细胞蛋白时,如果条件不当就会破坏两种相互作用蛋白形成的复合物的稳定性,使得免疫共沉淀实验失败。如果两种蛋白在细胞内的结合力确实非常弱,那么免疫共沉淀也难以成功。如果知道发生相互作用的两个蛋白都是胞核蛋白,那么可以通过提取核蛋白,再进行免疫共沉淀实验,这样会大大减低背景的干扰。关于两种蛋白质之间胞内的相互作用,有时确实无法用免疫共沉淀实验证明。
(1) 细胞内过表达蛋白的免疫共沉淀
证明蛋白质胞内相互作用时,可以选择一个高效瞬时过表达系统(至于这一系统是否有内
源性靶蛋白无关紧要)。一般采取 COS 细胞作为真核表达株。把两种蛋白基因共转染到 COS 细胞内进行表达。由于人为进行大量表达,所以在胞内两种蛋白形成相互作用的复合物的量也相应增多。如果你手头没有这两种蛋白的抗体,可以把这两种蛋白的一端分别加上标签以融合蛋白形式表达,然后用商业化的抗标签抗体进行免疫共沉淀和 Western blotting 检测。
(2) 细胞内源性蛋白的免疫共沉淀
把两种蛋白共转染到 COS 细胞内进行过表达,进行免疫共沉淀实验,相对容易成功,但是
这一结果毕竟具有人工性,不能代表生理条件下真实的蛋白质相互作用。要想克服这一弱点,
可以做内源性的免疫共沉淀实验。这一技术要求极高,难度极大,但也最有说服力。因为细胞内内源性蛋白含量低,结合在一起形成复合物的量更低,难以检测。首先要证明所选择的细胞系是否具有这两种内源性的蛋白。另外,用于免疫沉淀和 Western blotting 检测的抗体要好。细胞裂解、收集以及免疫沉淀时时条件要温和,以保持蛋白复合物的天然结构。
(3) 组织内蛋白的免疫共沉淀
在体外可以大量培养细胞,然后制备细胞提取物,做内源性免疫共沉淀。由此可以推广到做
组织内免疫共沉淀。取动物组织(脑、肝、脾等),切碎,匀浆,提取组织蛋白,进行免疫共沉淀实验。这一结果代表活体中蛋白质相互作用。
4, 蛋白质细胞内定位实验17-22
另一种经常用来检验蛋白质相互作用的方法是蛋白质细胞内定位技术。此法较为直观,可
以看到两种有相互作用的蛋白质在细胞内的分布(膜上、胞浆、胞核或其它细胞器等等)以及共定位的部位(在膜上共定位、在胞浆中某一部位或核内共定位等等)。有时相互作用的蛋白由于细胞内某种功能的需要结合在一起时,使得两种蛋白的分布发生变化。比如,某种蛋白也许在核内,当它与另一种具有穿梭功能的蛋白结合时,有可能被转运到胞浆中。这种情况的共定位则较为典型。在进行蛋白质共定位
(1) 利用有色荧光蛋白标记技术进行蛋白定位研究
此法也可称为活细胞定位。把两种具有相互作用的蛋白分别克隆到带有两种不同颜色荧光
蛋白(绿色荧光蛋白或红色荧光蛋白)的载体中,共转染到功能细胞中(一般选用 COS7 细胞)表达带有荧光的融合蛋白。这样,相互作用的两种蛋白就被标上不同的荧光,可以在细胞内用荧光显微镜直接观测。在进行精确细胞定位或共定位时,必须用共聚焦荧光显微镜观测。因为共聚焦荧光显微镜(相当于医院给病人诊断的 CT)观测的是细胞内一个切面上的颜色。如果在一个切面上在同一区域看到两种颜色,就提示这两种蛋白在该区域内有相互作用。普通荧光显微镜看到的是一个立体图象,无法确定蛋白质共定位现象。在进行定位或共定位同时,也可以对细胞核进行染色。这样,在细胞中就有三种颜色。细胞核的显色帮助你确定共定位发生的位置。上面介绍的活细胞定位,其优点是表达的荧光蛋白荧光强,没有背景,观测方便。但缺点
是相互作用的蛋白由于标上荧光蛋白,实际上是两个融合蛋白。融合蛋白的定位结果或共定位结果是否与天然蛋白分布一致,有待于进一步确定。而利用免疫荧光标记技术可以避免这一缺点。
(2) 利用双色或多色染色的免疫荧光技术进行蛋白定位研究
免疫荧光的原理是,首先把细胞进行固定,然后用待检测靶蛋白的抗体(一抗)与细胞内
靶蛋白进行免疫反应,再用荧光素(如 FITC 和 TRITC 等)标记的二抗与一抗进行反应。这样就在细胞内形成免疫复合物(靶蛋白----一抗---二抗),结果靶蛋白被标上颜色,然后可用共聚焦荧光显微镜观测定位与共定位结果。
免疫荧光技术最大优点就是可用来检测细胞内源性蛋白的定位及相互作用。当然也可以对
靶细胞进行转染表达目的蛋白,然后标记目的蛋白进行观测。免疫荧光技术的缺点是荧光相对较弱并且背景较高,结果受到干扰,所以这项技术不好掌握。为了结果的可靠性,要求严格设计阳性对照与阴性对照。
(3) 细胞内蛋白动态定位
有时细胞在正常状态下,有相互作用的蛋白在胞内可能暂时分开,没有共定位现象发生。
但是在某一个特定情况下,如细胞受到外界刺激时,细胞本身会产生应急反应,这时暂时分离的蛋白有可能发生相互作用,并产生共定位现象。所以在进行共定位研究时,可根据具体情况具体分析,必要时观测细胞内蛋白动态定位结果。