中科院:蜡样芽孢杆菌降解孔雀石绿?蛋白组学分析机制
导读
孔雀石绿(MG)作为一种染料的广泛应用引起了人们的广泛关注。蜡样芽孢杆菌能对抗MG的毒性作用,并能有效地对MG进行脱色。然而,基于蛋白质组学数据的关于其潜在适应和降解机制的详细信息却很少。本研究采用等压相对和绝对定量标记(iTRAQ)辅助定量方法,分析了蜡样芽孢杆菌降解MG的分子机制。在此基础上,在MG生物降解过程中,我们鉴定出209个上调蛋白和198个下调蛋白,错误发现率不超过1%。基因本体论和KEGG分析表明,差异表达的蛋白质在代谢过程、催化活性、抗氧化活性和对刺激的反应等方面都有丰富的表达。此外,利用实时qPCR进一步证实了参与苯甲酸降解的调控蛋白。参与苯甲酸降解途径的蛋白质BCE_4076(乙酰辅酶A乙酰转移酶)、BCE_5143(乙酰辅酶A乙酰转移酶)、BCE_5144(3-羟酰基辅酶A脱氢酶)、BCE_4651(烯醇辅酶A水合酶)和BCE_5474(3-羟酰基辅酶A脱氢酶)可能在蜡样芽孢杆菌对MG的生物降解中起重要作用。这项研究的结果不仅提供了蜡样芽孢杆菌在MG负荷下蛋白质组学变化的综合观点,而且还揭示了蜡样芽孢杆菌降解MG的机制。
论文ID
原名:iTRAQ-facilitated proteomic analysis of Bacillus cereus via degradation of malachite green
译名:iTRAQ促进蜡样芽孢杆菌降解孔雀石绿的蛋白质组学分析
期刊:Journal of Microbiology
IF:3.422
发表时间:2021.02
通讯作者:余志晟
通讯作者单位:中国科学院大学
实验设计
1. 首先评估了不同浓度MG存在下蜡样芽胞杆菌的生长速度;
2. iTRAQ蛋白质组学方法定量分析被用于鉴定蜡样芽孢杆菌降解MG所涉及的蛋白质;
3. 利用MG胁迫下下调和上调的蛋白质进行了KEGG途径分析;
4. 分析参与碳代谢和能量代谢的蛋白质、参与核苷酸修复和应激反应的蛋白质分析、谷胱甘肽代谢相关的蛋白至、苯甲酸酯降解过程中的蛋白质。
实验结果
1. 生长速度和脱色效率
为了研究蜡样芽胞杆菌生长速率的变化如何反映MG负荷反应,我们评估了不同浓度MG存在下蜡样芽胞杆菌的生长。图1总结了蜡样芽胞杆菌在50、200和400 mg/LMG存在下的CFU和细胞生长曲线。用不同浓度的MG培养蜡样芽胞杆菌表明,在低于400 mg/L的MG浓度下,蜡样芽胞杆菌的菌落数达到106 CFU/mL,我们在这些细菌细胞之间没有生长差异,如图1A所示。然而,在400 mg/L的MG浓度下,蜡样芽胞杆菌的菌落数达到104 CFU/mL。此外,蜡样芽胞杆菌在不同MG浓度下的生长曲线进一步证实了菌落计数结果(图1B)。这些结果表明,高浓度MG对蜡样芽孢杆菌的生长有抑制作用。MG在许多微生物中的生长抑制已被证实,例如金黄色葡萄球菌、黄孢原毛平革菌、枯草芽孢杆菌和酿酒酵母。MG的抑制作用归因于抑制细菌DNA复制和增强细胞中的活性氧物种。在本研究中,400 mg/L的MG对蜡样芽孢杆菌的生长有明显的抑制作用。因此,虽然MG在低浓度下对蜡样芽孢杆菌的生长没有抑制作用,但为了维持正常生长,细菌受到了MG的遗传毒性和细胞毒性的胁迫和损伤。由于比较同一培养阶段的细菌蛋白质组很重要,因此我们选择了对蜡样芽孢杆菌生长无抑制作用的MG浓度(200 mg/L),并通过离心在对数后期收集所有用于iTRAQ分析的蛋白质样品。
我们分析了蜡样芽孢杆菌在不同MG浓度培养基中的脱色效率。尽管当染料浓度增加到400 mg/L时,染料的总脱色量急剧下降(图1C),但随着染料初始浓度的增加,MG的数量增加,Chen等人也报告了初始MG浓度增加时,去除的颜色量增加。这可能是由于较高的染料初始浓度可以增强驱动力,克服染料在细胞与MG之间的传质阻力。
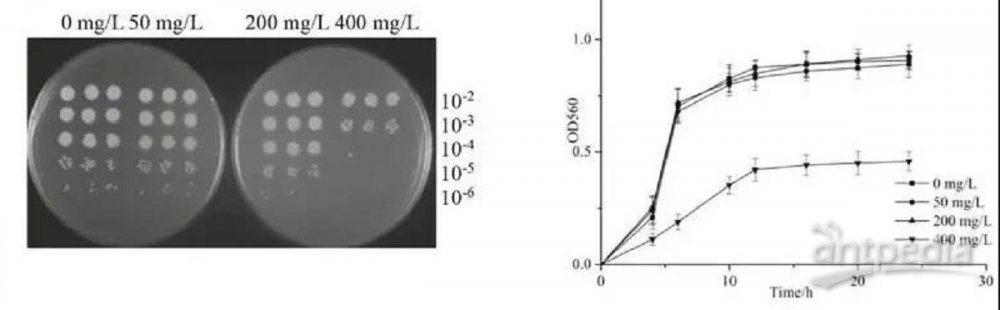
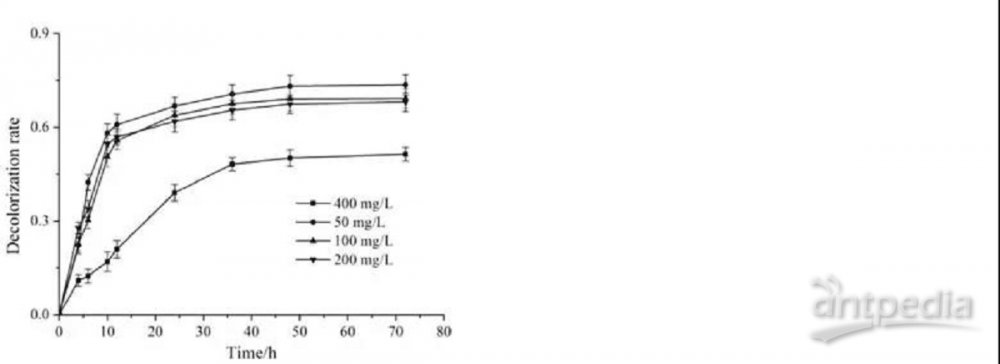
图1 蜡样芽孢杆菌在MG负荷下的生长及脱色效果
(A)蜡样芽孢杆菌在LB琼脂平板上不同MG浓度培养后的CFU谱。(B)蜡样芽孢杆菌的生长曲线。(C)不同初始染料浓度对蜡样芽孢杆菌脱色的影响。
2. 蜡样芽孢杆菌iTRAQ定量蛋白功能分类研究
在这项研究中,iTRAQ蛋白质组学方法被用于鉴定蜡样芽孢杆菌降解MG所涉及的蛋白质反应。皮尔逊相关性用于验证生物复制品的分析再现性。我们在整个检测范围内获得了一条变化很小的45度对角线,表明实验具有高度的可重复性(参见补充数据图S2)。在生物信息学分析中,我们将蛋白得分为1.3或更高作为阈值,从1725个独特肽中鉴定出了407个差异调控蛋白(见补充数据表S2)。由iTRAQ蛋白质组学鉴定的蛋白质的长度和肽数的分布以及序列覆盖率显示在补充数据图S3AC中。
407个差异表达蛋白中,209个表达上调,198个表达下调(iTRAQ比值1.5或0.667,P值均<0.05)。根据GO分析,这些调节蛋白根据细胞成分、生物过程和分子功能进行功能聚类,如图2A C所示。在三个主要类别中,细胞成分类别的最大比例由细胞部分(72.3%)和大分子复合物(17%)表示;在生物过程类,代谢过程(70.8%)和细胞过程(17.3%);分子功能类,蛋白质的催化活性(65.4%)和结合活性(19.1%)较高(图2)。大量参与代谢过程和催化活性的蛋白质受到不同程度的上调或下调。此外,参与抗氧化活性和刺激反应的蛋白质均上调(图2)。特别是,GO类代谢过程在MG负载样品中包含33种上调的蛋白质物种,包括合成酶(PdxS、CarA、CarB、RibH、ArgJ、ArgG和MiaB)、连接酶(SucC、SucD、ArgS和GltX)以及H+转运ATP酶亚基ε和γ。此外,MG负载样品中五种脱氢酶(GuaB、Mdh、GcvPB、PdhA和RocA)的催化活性也增加,而三种脱氢酶(MtnB、IlvD和HisD)则降低。在抗氧化活性方面,谷胱甘肽过氧化物酶(BsaA)、有机过氧化氢抗性蛋白(BCE_)、烷基过氧化氢还原酶(AhpC和AhpF)表达上调3~7倍。在对刺激的反应中,黄血球蛋白(Hmp)、丝氨酸羟甲基转移酶(GlyA)、ATP依赖性解旋酶/核酸酶亚单位A(AddA)和DNA错配修复蛋白(MutS)等4种蛋白均增加。因此,MG负荷通过影响细胞生理的许多方面,包括新陈代谢、生理活动和应激反应,强烈地重塑了蜡样芽孢杆菌的蛋白质组。
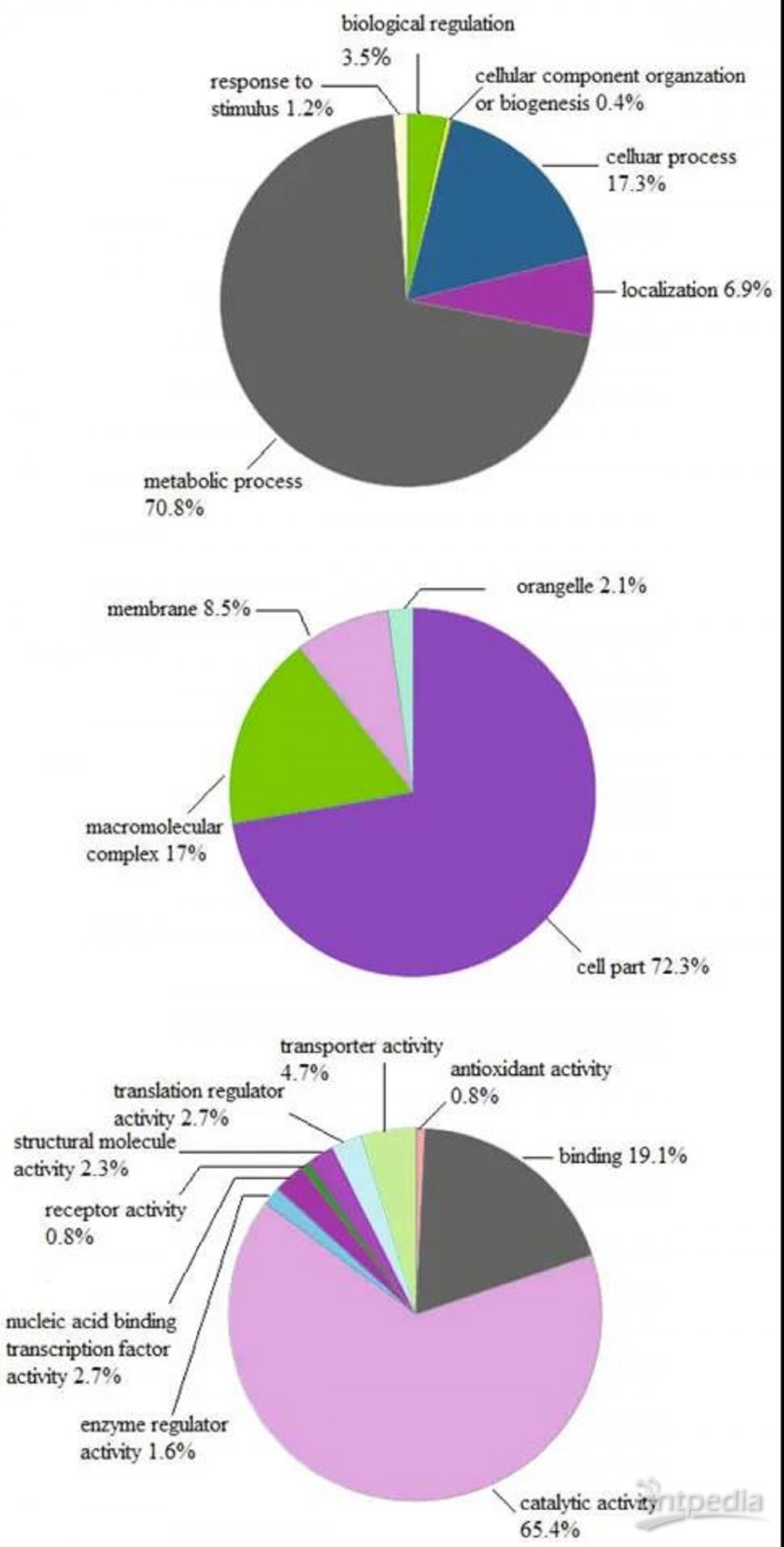
图2 MG负荷下蜡样芽孢杆菌差异表达蛋白的基因本体分析
(A)生物过程类的改变蛋白质;(B)细胞成分类的改变蛋白质;(C)分子功能类的改变蛋白质。
3. 通径分析
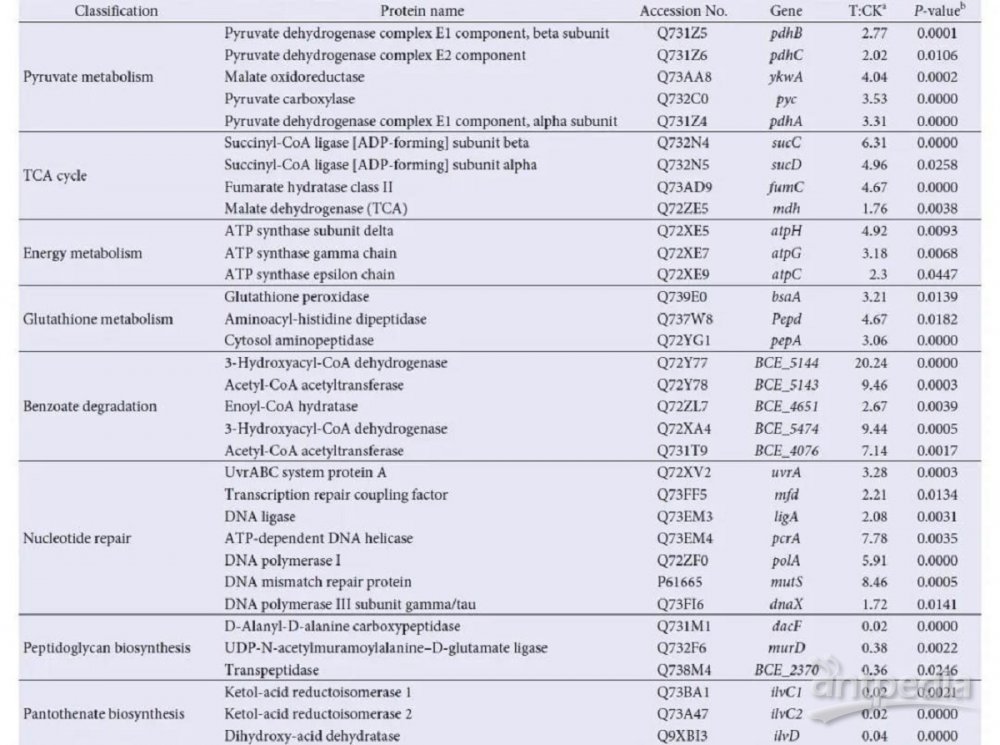
为了更好地理解MG胁迫对细菌途径的影响,我们利用MG胁迫下下调和上调的蛋白质进行了KEGG途径分析(图3)。在MG负荷下,45种蛋白与碳代谢有关,如糖酵解/糖异生、三羧酸循环(TCA)和丙酮酸代谢(表1)。相比之下,在MG处理的蜡样芽孢杆菌细胞中,有13种下调的蛋白参与氨基酸代谢和生物合成,包括缬氨酸、亮氨酸、精氨酸、脯氨酸和异亮氨酸代谢。此外,参与其他生物过程的蛋白(如脂肪酸代谢、肽聚糖生物合成和泛酸生物合成(表1))被下调。这些结果表明,MG以细胞应激的形式暴露,可以影响蜡样芽胞杆菌的多种代谢途径,这可能解释了高浓度MG对蜡样芽胞杆菌生长的抑制作用。值得注意的是,有5种上调蛋白(UvrA、Mfd、LigA、PcrA和PolA)与核苷酸切除修复有关,表明MG负载后可能存在DNA损伤,与DNA修复功能相关的蛋白在MG生物降解中起作用。因此,DNA修复蛋白的上调可能是蜡样芽孢杆菌抵御MG负荷的一种保护机制,而MG胁迫下特异性代谢流量的降低可能是一种综合性的抗MG策略。

图3 基于KEGG富集分析的MG负荷下蜡样芽孢杆菌差异表达蛋白的途径分类
丰富因子,差异表达蛋白数量占相关通路总基因数量的比例。有超过5种蛋白质变化的途径被显示。
表1 部分差异表达蛋白及其在选定功能类别中的指定或假定功能
4. 参与碳代谢和能量代谢的蛋白质分析
与丙酮酸代谢和TCA相关的蛋白质在MG存在下在蜡样芽孢杆菌中表现出增加的表达;这些蛋白质包括二氢硫酰脱氢酶、丙酮酸脱氢酶复合物和乙酰辅酶A合成酶。此外,丙酮酸脱氢酶复合物(PDHC)是一种参与能量代谢的重要酶,在MG处理的样品中表现出明显的上调。PDHC是TCA的底物,它催化丙酮酸生成乙酰辅酶A。这一过程在从糖酵解到TCA的过程中起着至关重要的作用。此外,果糖1,6-二磷酸酶和丙酮酸羧化酶的增加表明糖异生作用增加。因此,这些结果表明,在MG胁迫下,参与碳代谢的蛋白质表达增加可能会增加碳骨架的产生或间接增加能量供应。此外,参与能量代谢的ATP合酶复合物的三个亚基ATP合酶β亚基(AtpC)和ATP合酶γ链(AtpH和AtpG)显著上调。这些发现与观察到的蜡样芽孢杆菌在MG降解过程中碳水化合物代谢途径成员的上调是一致的。Yi等人发现,在大肠杆菌工程菌株的三苯基锡生物降解过程中,ATP合成酶的三个亚基(AtpA、AtpD和AtpG)的表达显著上调。此外,Szewczyk等人认为,能量相关蛋白表达的增加似乎是外源生物降解过程中的普遍结果。在本研究中,这三个ATP合酶亚基的上调表达表明蜡样芽孢杆菌在MG生物降解过程中需要大量的能量。
5. 参与核苷酸修复和应激反应的蛋白质分析
在上述改变的蛋白中,许多核苷酸切除修复蛋白的表达增加。这可能是由于细胞内的代偿机制,因为MG可以结合几乎所有可用的DNA位点,并导致细菌DNA损伤。尤其是参与核苷酸切除修复和错配修复的7种蛋白(UvrA、Mfd、LigA、PcrA、MutS、PolA和DnaX)上调3-8倍。其中,UvrA催化DNA损伤的识别和处理。LigA催化双链DNA中5-磷酰基和3-羟基之间磷酸二酯键的形成,对DNA复制和受损DNA修复至关重要。DnaX负责大多数复制DNA合成;黄素血蛋白(Hmp)在对各种有毒氮化合物的诱导反应中起着核心作用。我们发现MG负荷组上调了1.82倍。此前,有报道称,丝氨酸羟甲基转移酶(GlyA)可提高大肠杆菌对高渗胁迫的耐受性,在200 mg/L的MG负荷组中,它被发现分别上调了2.21倍。值得注意的是,参与肽聚糖生物合成的三种蛋白质(DacF、MurD和BCE_2370)表达下调。肽聚糖在细菌中起结构作用,并抵消渗透压。因此,在蜡样芽孢杆菌中,高MG浓度和低肽聚糖生物合成可能会引起细胞的渗透胁迫,而上调GlyA可能是与MG相似的压力反应策略。
6. 谷胱甘肽代谢相关蛋白的分析
纺织染料作为环境污染物在细胞氧化应激的产生中起着关键作用,并阻碍微生物的生长。结果表明,在蜡样芽孢杆菌降解MG的过程中,谷胱甘肽过氧化物酶、氨酰组氨酸二肽酶、胞浆氨肽酶等与谷胱甘肽代谢有关的抗氧化酶表达上调。GSH是生物体内一种重要的抗氧化剂。它能清除多余的活性氧自由基,减轻细胞代谢过程中膜脂过氧化造成的损伤。先前的研究表明,GSH在降低MG的毒性作用中起着关键作用。在最近的一项研究中,Yildirim等人还证实,蚤状钩虾中GSH、超氧化物歧化酶、谷胱甘肽S-转移酶和谷胱甘肽过氧化物酶水平的增加提高了云芝对MG的脱色能力。这些发现表明细胞需要活性抗氧化蛋白来抵抗MG的毒性。GSH代谢相关蛋白的上调可能增强蜡样芽孢杆菌抵御氧化应激和促进MG降解的能力。
7. 苯甲酸酯降解过程中的蛋白质分析
有两种重要的机制被证明介导了三苯甲烷染料的降解。蜡样芽孢杆菌能将MG降解为4,4-二(二甲氨基)二苯甲酮和另一种中间代谢物苯酚。这一过程类似于微球菌BD15对MG的降解。在微球菌菌株BD15中,4,4-双(二甲氨基)-二苯甲酮被降解为N-二甲基苯胺和4-二甲氨基苯甲酸,然后进一步转化为其他化合物进行降解。在这项研究中,与苯甲酸降解相关的蛋白质(BCE_5474、BCE_5144、BCE_4651、BCE_4076和BCE_5143)在蜡样芽孢杆菌的MG生物降解过程中显著上调。我们用RT-qPCR检测200 mg/L MG胁迫下BCE_4651、BCE_5143、BCE_5144、BCE_4076和BCE_5474基因的表达水平(图4A),并且这些结果与蛋白质组学结果一致。两种分析都表明,这些基因的转录和翻译水平在蜡样芽孢杆菌降解MG的过程中显著增加。因此,我们假设4,4-二(二甲氨基)二苯甲酮在蜡样芽孢杆菌中转化为苯甲酸盐进一步降解。之后,苯甲酸降解的最终产物通过TCA进一步降解(图4B)。Zhang等人也报道了苯甲酸盐降解途径富集在具有复杂苯环结构的染料活性黑5的降解中。基于上述蛋白质的数量和生物学功能,我们的研究结果表明苯甲酸降解途径可能在蜡样芽孢杆菌降解MG的过程中起重要作用。
基于目前的蛋白质组学研究结果和以往的文献,我们提出了蜡样芽孢杆菌对MG的潜在降解途径,如图5所示。蜡样芽孢杆菌生物降解MG的初始步骤已在不同的研究中得到验证。MG被氧化分解成4,4-二(二甲氨基)二苯甲酮和苯酚。芳香族聚合物降解的第二步是对芳香族化合物单体的分解代谢。然而,参与这些过程的蛋白质很少上调,这可能与该菌株对MG的固有高度脱色有关,因为蜡样芽孢杆菌的粗酶具有降解MG的能力。在MG处理中,上调的主要蛋白参与了苯甲酸盐降解途径和TCA途径。已提出不同菌株如假单胞菌、伯克霍尔德菌参与MG降解的TCA途径。这项研究表明,苯甲酸降解途径可能是负责进一步降解芳香化合物从MG。因此,本研究提出的蜡样芽孢杆菌对MG的降解途径为研究MG的生物降解机理提供了新的思路。

图4 苯甲酸降解途径在MG的降解过程中起着重要作用
(A)MG负荷下蜡样芽孢杆菌苯甲酸降解相关基因的转录水平。结果代表三次实验的平均值。误差线表示一个标准差。(B)KEGG数据库中显示了从苯甲酸降解到柠檬酸循环的代谢流路径模型。
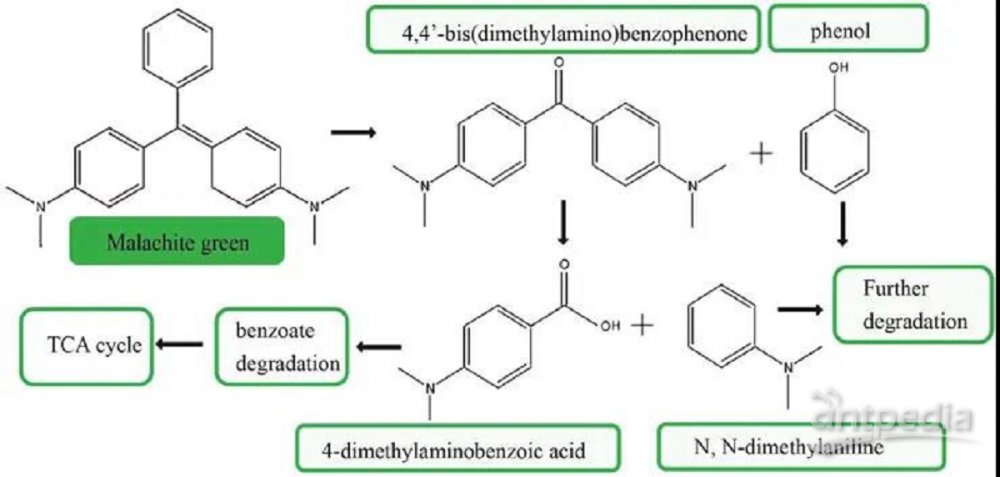
图5 根据蛋白质组学的研究结果和前人的研究成果所提出的蜡样芽孢杆菌降解MG途径
结论
总之,我们的研究在蜡样芽胞杆菌中发现了一大类差异表达蛋白,这些差异表达蛋白参与了使用基于iTRAQ的LC-MS/MS降解MG的过程。这些差异表达蛋白参与了MG胁迫的保护、促进细菌生长以及在MG生物降解过程中最小化氧化应激。此外,我们发现在MG的降解过程中苯甲酸盐的降解途径增强。然而,需要进一步的研究来构建敲除突变体来验证MG降解相关的基因功能。这些结果揭示了MG生物降解的第一个全面的蛋白质组,为MG生物降解的分子机制提供了新的见解。