转座酶的因子类型
插入序列
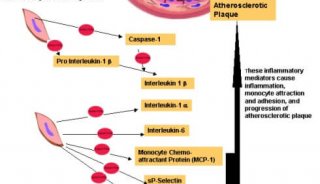
IS1是基因组中可移动的遗传因子家族中的成员之一,它可以整合到宿主非同源位点上,这就是转座(tronsposition),若IS插入到某基因内,通常这个基因就会失活。这种精确的作用取决于IS有关的状况。例如IS2因子以同一方向插入到染色体中,则会减少基因的表达,但以相反方向整合,则会增加基因表达。相比这下IS1因子不论以什么方向插入都会降低基因的表达。
IS原来分为IS1~4,后面的数字反应它们被分离的先后次序和转座因子的总数无关。
正常的细菌染色体和质粒都含有IS。E.coli的标准品系似乎含有各种拷贝数(<10)的IS因子。在原核生物中IS家族有很多成员,它们的结构相似,两端都有短的正向重复序列(directrepeats,DR)(靶序列),略长的反向重复序列(invertedrepeats,IR)以及1kb左右的编码区,它仅编码和转座有关的转座酶。对靶的选择有三种形式:随机选择,热点选择和特异位点的选择。
由于IS中有反向重复序列的存在,因此如果质粒上有IS,那么经变性和复性后,在电镜下可以观察到茎环结构的存在(图23-30)。
IS的转座是由转座酶(transposase)催化的,它由IS编码。首先转座酶交错切开宿主靶位点,然后IS插入,与宿主的单链末端相连接,余下的缺口由DNA聚合酶和连接酶加以填补,结果使插入的IS两端形成了DR或靶重复。
类插入序列
(IS-likeelements)是指IS10R,IR50R和IS903,它们的结构和IS相似,但不独立存在。而是作为复合转座子(compositetransposons)两臂的组件。
复合转座子
复合转座子要比IS长得多,中心区域编码抗性标记。不同的复合转座子的抗性标记不同。复合转座子两端的组件由IS和类IS组成。有的二侧组件相同(如Tn903),有的不同(如Tn10)。有的方向相同(如Tn9),有的方向相反(如Tn903,10,5)。有的皆有功能(如Tn903,10),有的仅右侧组件有功能。
据推测当两个独立的组件与中心区域连接时发展成复合转座子。这个结构可能由IS因子转座到紧靠供体位点的受体位点上。这两个组件可能相同或存在差异。单个组件转座整个复合转座子的能力解释了两个组件保持活性不受选择压力的影响。
在复合转座中是什么替代单个组件而担负起座的作用呢?特别是在两个组件都有功能的情况下。在Tn9例子中,它的组件是IS1因子,推测它们都有功能。为什么转座是整个复合转座,而不是IS的本身呢?
两个IS因子能使夹在其中的任何序列转座。如一个环状复制子上的Tn10它的两个组件IS10L和IS10R,也可以看成是复制子序列两端的组件,因此在进一步转座时它不仅可以使Tn10转座,也可心使复制子序列转座,但两侧面组件的序列与在Tn10中不同。
TnA家族
转座子(transposon,Tn)另一家族是TnA,长约5kb左右,两端具有IR,而不是IS,中部的编码区不仅编码抗性标记,还编码转座酶和解离酶。
TnA是复制转座的转座子。在此家族中Tn3和Tn1000(γδ)研究得最深入。通常末端有38bp左右的IR,在两个IR中任一个顺式作用缺失都会阻止转座。5bp的正向重复是靶位点产生的。它们都带有抗性标记(图23-33)。
TnpA介导转座分为两步,分别由tnpA,、tnpR编码的转座酶(transposase)和解离酶(resolvase)来完成的。这两个酶是通过实变鉴别出来的。此类转座和末端的IR有关。解离顺序(res)是内部的特殊位点,只有TnA家族才具有这一位点。
tnpA突变体是不能转座的,这个基因的产物是一种转座酶。它可以结合在末端38bpIR中的25bp的序列上。E.coliIHF的结合位点就在转座酶结合位点附近(在靶位点附近)。而转座酶结合和IHF的结合是协同性的。转座酶识别末端重复序列,并可以使靶DNA产生5bp的交错切割使转座子插入。IHF是一种DNA结合蛋,常在E.coli中组装成一个大分子,它在转座反应中并不起作用。
tnpR的产物具有双重功能,一是作为基因表达的阻遏物,另一个作用是具有解离酶的功能。tnpR突变将增加转座预率,这是由于TnpR阻遏了tnpA和它自己的基因转录,TnpR蛋白失活就使TnpA合成增多,结果增加了转座的频率。这表明TnpA转录酶的数量一定是转座的限制因子。
tnpA和tnpR基因之间有一个富含A-T的内部顺式控制区,TnpR的两种功能就是通过和此区的结合来实现的。