







Effect of ethanol on development of Danio reriro
Objective
The objective of the experiment is to determine the effects of ethanol exposure on the embryonic development of zebrafish through observation of physical defomities.
Embryonic development of zebrafish, Danio reriro, is affected by ethanol in a manner similar to higher vertebrates (Blader and Strähle, 1998). Therefore, investigating the effects of ethanol on zebrafish embryo many promote knowledge and understanding of fetal alcohol syndrome in humans. Blader and Strahle confirmed that exposure of zebrafish embryos to ethanol causes cyclopia and craniofacial abnormalities and alters gene expression in the ventral aspects of the fore and midbrain (1998). Exposure of zebrafish embryos to ethanol also induces developmental abnormalities of the notochord and spinal cord, and malformation of the body trunk (Blader and Strähle, 1998).
In order to understand the origin of the deformities, it is necessary to know how embryonic cells of zebrafish normally migrate during gatrulation. During gastrulation, blastoderm cells epibolize over the yolk. When about half the yolk cell becomes covered with blastoderm cells, the margin of epibolizing blastoderm cells form the epiblast and the hypoblast. Epiblast and hypoblast intercalate to form the organizer, known as the embryonic shield, at the dorsal side of the embryo (Gilbert, 2003). Hypoblast cells of the embryonic shield converge, extend anteriorly, and narrow along the dorsal midline of the hypoblast (Gilbert, 2003). Some of these cells eventually form the prechordal plate and the notochord (Gilbert, 2003). Prechodal plate cells seem to be crucial in explaining the origin of ethanol induced deformities.
Though little is known about the underlying mechanisms behind the deformities, it is known that the ectopic position of the prechordal plate cells is characteristic of zebrafish exposed to ethanol (Blader and Strähle, 1998). It is, therefore, believed that ethanol is responsible for the abnormal migration of prechordal plate cells that ultimately causes cyclopia and other deformities. The prechordal plate cells express genes like goosecoid and islet-1, which control cell differentiation in the anterior region of the embryo (Blader and Strähle, 1998). Hence, prechordal-specific genes are expressed ectopically to bring about the observed deformities.
In addition to the deformities, ethanol appears to cause abnormal cell death (Sulik et al., 1988). Ethanol exposure at early development stages results in significant death among the cells destined to give rise to facial structures (Sulik et al., 1988). Ethanol seems to achieve apoptosis by activating the cells’ self-destruction machineries (Sulik et al., 1988). Therefore, in addition to the ectopic prechordal plate-specific gene expression, ethanol induced apoptosis appears to contribute to the observed deformities in zebrafish embryos (Sulik et al., 1988).
This experiment seeks to determine the effects of exposure of ethanol on zebrafish development through observation of physical deformities. Most of the procedures were adopted from Blader and Strähle’s experiment (1998). 30% epiboly stage, also called the late blastula stage, is followed immediately by gastrulation (Gilbert, 2003). Therefore, embryos at 30% epiboly stage at the latest will be used to ensure that ethanol will affect cell migration before it occurs. The results will be compared to Blader and Strahle’s.
Materials
Siphon
Zebrafish Embryo Solution
Fine fry net or mesh filter
ethanol
large-bore glass pasteur pipets
60mm glass Petri dishes (8)
dissecting microscope
zebrafish embryos at dome/30% epiboly stage (40)
Incubator, 28°C
A. Isolation of Zebrafish Embryos
1. The night before embryo collection, vacuum the tank well with a siphon to remove debris several hours after the last feeding.
2. Deposit a layer of washed marbles to cover the entire bottom surface of the tank.
3. Do not disturb for the first hour after dawn to allow fish to mate.
4. Collect embryos and fertilized eggs from between the marbles with a siphon then and collect with a fine fry net or mesh filter.
5. Transfer the net to a glass dish full of tank water and examine the embryos with a dissecting microscope at low magnification.
6. Transfer healthy embryos to a clean glass Petri dish containing embryo media with large-bore glass Pasteur pipettes. Discard any that are cloudy or ruptured.
7. When not being used for the experiment, the embryos should be kept at 28°C (Zebrafish embryos’ rapid development makes constant temperature crucial for obtaining a reproducible group of embryos for experimentation).
*protocol adopted from DB lab Website: Isolation of Zebrafish Embryos
B. Ethanol Treatment
1. Select 40 healthy zebrafish embryos that are at 30% epiboly stage (Figure 1 below).
2. Prepare four separate petri dishes that contain each of the following four: normal
zebrafish embryo medium, 1%, 2%, and 3% ethanol zebrafish embryo medium
solutions.
3. Take embryos out of the incubator (set at 28°C) and place 10 zebrafish embryos
into each petri dish. Place all four dishes in the incubator. Then wait for 3 hours
for the ethanol to take effect.
4. After the 3 hours, remove the dishes from the incubator and move the embryos
into new petri dishes containing normal zebrafish embryo solution. Different
batches of embryos must go into different dishes. Place the dishes in the
incubator and wait for 24 hours.
5. Return after 24 hours. Observe the embryos for abnormalities and photograph
them.
6. Return 24 hours later to observe the embryos for abnormalities. Photograph them.
*protocol adapted from: The Effects of Ethanol on Zebrafish Development: Jeff Mindel, F&M.
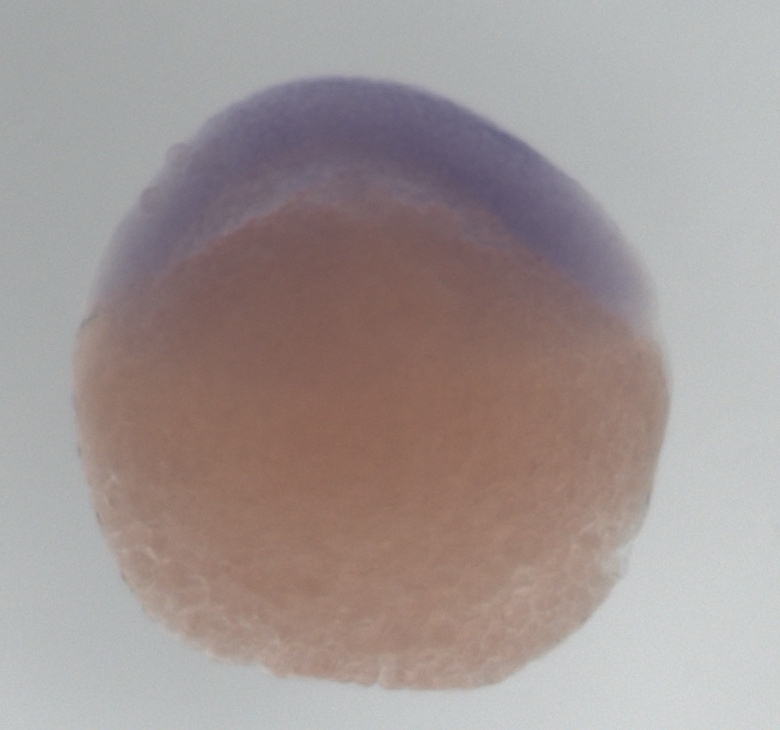
Figure 1. Zebrafish embryo at 30% epiboly stage (4 3/2 h). Top is the animal pole.
24 hours after the exposure of ethanol, cyclopia was observed among the embryos cultured at 3% ethanol (Figure 1, Table 1). No cyclopia was observed in embryos at 1% or 2% ethanol solutions. For both concentrations, eyes developed normally; the vesicles were darkly pigmented and the eyes were normally spaced (Figures 1B, 1C). At 3%, 1 cyclopic embryo was observed. The rest of the surviving embryos showed no signs of cyclopia. The cyclopic embryo’s eyes were fused and were deficient in pigmentation (Figure 1D). In addition to cyclopia, embryos exposed to 3% ethanol showed varying degrees of impaired development of posterior structures, more specifically of the trunk and tail, 24 hours after their exposure to ethanol (Figure 3). Again, no deformities were observed in the posterior regions of those at 1% and 2% ethanol solutions (Figures 3B, 3C). 2 embryos, 1 of whom was the aforementioned cyclops, cultured in 3% ethanol solution displayed malformation of the trunk and tail (Figures 3D, 3E). The non-cyclopic embryo (eyes were not fused but had shortened distance between them) with posterior deformities had enlarged allantois with a shortened tail (Figure 3D). Moreover, much of the pigmentations normally observed along the dorsal side of untreated embryos were absent (Figure 3D). The cyclopic embryo displayed similar symptoms with increased severity (Figure 3E). At the 48 hour stage, there were no posterior deformities observed among embryos cultured at 1% and 2% ethanol solution (Figure 4, Table 1). All surviving embryos grown at 1% and 2% solution that hatched displayed normal swimming movements. At 3%, the cyclopic embryos with deformed posterior structures continued to develop with the aforementioned deformities which, at the 48 hour stage, ultimately rendered the embryos immobile. The non-cyclopic embryo with defective trunk and tail died. The cyclopic embryo failed to hatch, and when artificially pulled out of the shell, did not show normal swimming motions. In fact, the embryo was completely immobile due to the severity of its trunk and tail deformites. Figure 1. Ethanol treatment causes cyclopia at 3%. Zebrafish embryos treated at the dome/30% epiboly stage with 1%,2%, and 3% ethanol- Zebrafish Embryo Medium (ZEM) solution for 3 h and cultured in ZEM for 24 h in the absence of ethanol. No cyclopia was observed in 1% and 2%. 1 cyclopic embryo was observed out of 10 at 3%. (A) Untreated control embryo. (B) 1% ethanol treated embryo. (C) 2% ethanol treated embryo. (D) 3% ethanol with cyclopia. Eye vesicles are fused along the midline of the head. Arrows indicate eyes. Click on the picture for enlarged version. Figure. 2. Ethanol treatment causes cyclopia at 3%. Embryos treated at the dome/30% epiboly stage with increasing concentration of ethanol for 3 h and cultured for 48 h in Zebrafish Embryo Medium in the absence of ethanol. Cyclopia observed at 3% only. (A) Untreated control embryo. (B) 1% ethanol treated embryo. (C) 2% ethanol treated embryo. (D) 3% ethanol with cyclopia. Eye vesicles are fused along the midline of the head. Embryos are shown in frontal views, dorsal up. Arrows indicate eyes. Click on the picture for enlarged version. FIG. 3. Ethanol treatment impairs trunk and tail development at 3%. Embryos were treated from the dome/30% epiboly stage with increasing concentration of ethanol for 3 h and then cultured for 24 hours in the Zebrafish Embryo Medium in the absence of ethanol. (A) Untreated control embryo. (B) 1% ethanol treated embryo. (C) 2% ethanol treated embryo. (D) Non-cyclopic embryo generated by 3% ethanol treatment showing mild deformities of the trunk and tail. (E) Cyclopic embryo generated by 3% ethanol treatment showing severe trunk and tail deformities. Arrows indicate defective trunk and tail. FIG. 4. Ethanol treatment impairs trunk and tail development at 3%. Embryos were treated from the dome/30% epiboly stage with increasing concentration of ethanol for 3 h and then cultured for 48 hours in the Zebrafish Embryo Medium in the absence of ethanol. (A) Untreated control embryo. (B) 1% ethanol treated embryo. (C) 2% ethanol treated embryo. (D) Cyclopic embryo generated by 3% ethanol treatment showing severe trunk and tail deformities. Arrows indicate trunk and tail deformities. Table 1. Developmental abnormalities induced by ethanol exposure. Embryos were treated from the dome/30% epiboly stage with 1%, 2%, and 3% ethanol- Zebrafish Embryo Medium (ZEM) solution for 3 h and cultured in the ZEM for a total of 48h in the absence of ethanol. 10 embryos were subjected to each concentration. Cultures were observed twice: 24 h after the treatment and 48 h after the treatment. Dead embryos with deformities were not counted. *One of the 2 also had cyclopia, the same embryo denoted by 1 in the 24 cyclopia column. Table 2. Viability of zebrafish embryos exposed to different concentration of ethanol concentrations at two 24 h intervals. Embryos were treated from the dome/30% epiboly stage with with 1%, 2%, and 3% ethanol- Zebrafish Embryo Medium (ZEM) solution for 3 h and cultured in the ZEM for a total of 48h in the absence of ethanol. 10 embryos were exposed to each concentration. Cultures were checked twice: 24 h after the treatment and 48 h after the treatment.
48 hours after the ethanol exposure, no cyclopia was observed in embryos grown at 1% and 2% ethanol solution (Figure 2, Table 1). For both solutions, eye development took place normally (Figures 2B, 2C). At 3%, the cyclops observed at the 24 hour stage advanced with its abnormal eye development as evidenced in dark optical pigmentation not seen previously (Figure 2D).
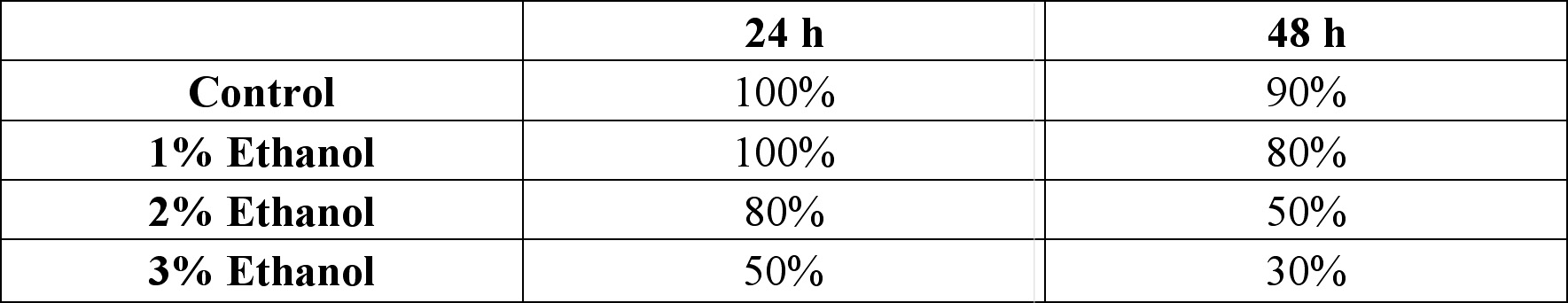
Another notable observation made was the viability of the zebrafish embryos (Table 2). 24 hours after the ethanol exposure, it was observed that increasing of ethanol concentration correlated with increasing death. There was no death among embryos culture in control and 1% ethanol solution. However, there were 2 deaths in 2% solution and 5 in 3% solution. When checked 48 hours after the ethanol exposure, viability of the embryos in each concentration was as following: 90% in control, 80% in 1%, 50% in 2%, and 30% in 3% (Table 2).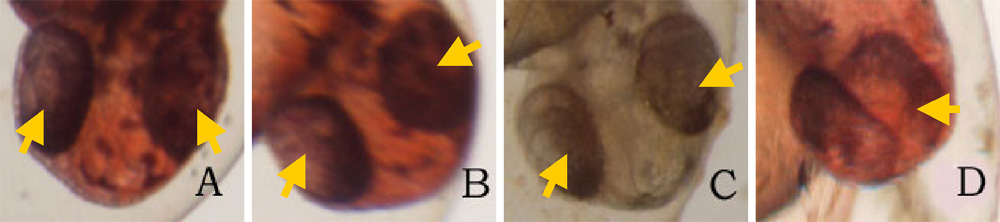
Click on the picture to view the enlarged version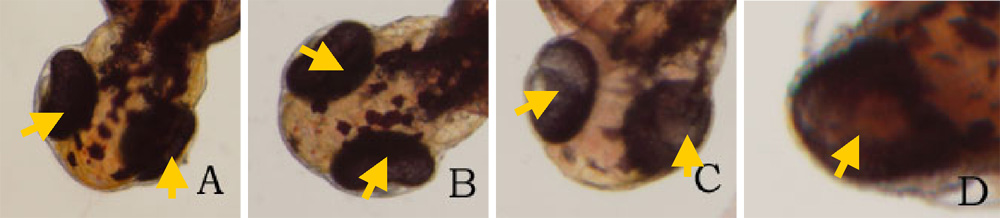 Click on the picture to view the enlarged version
Click on the picture to view the enlarged version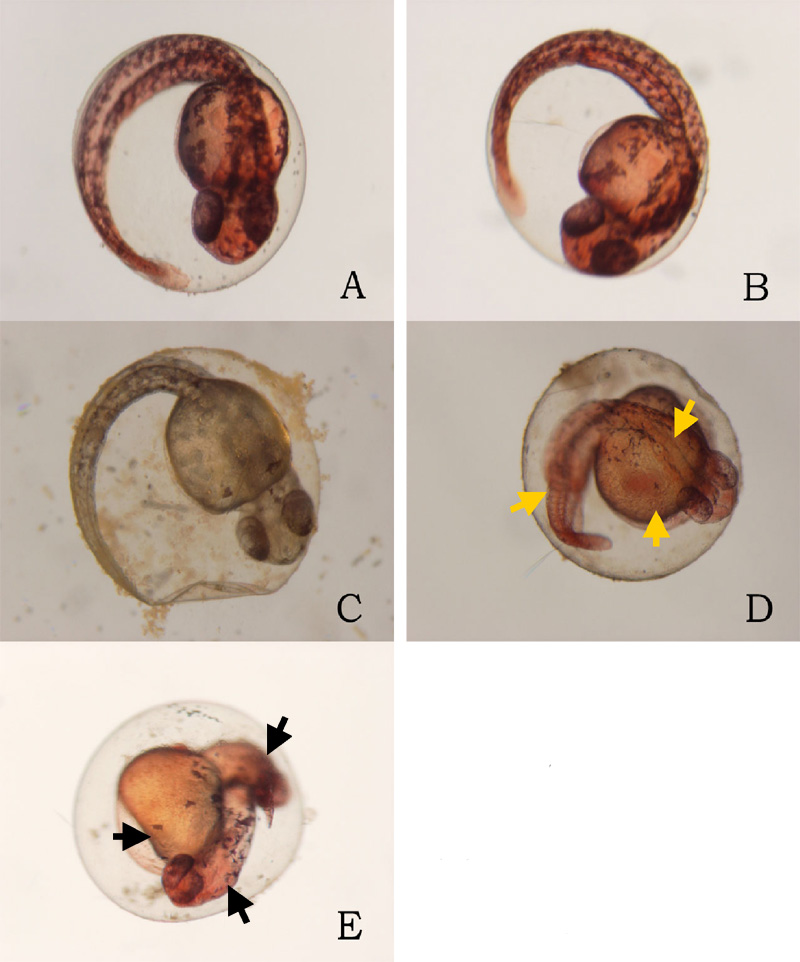

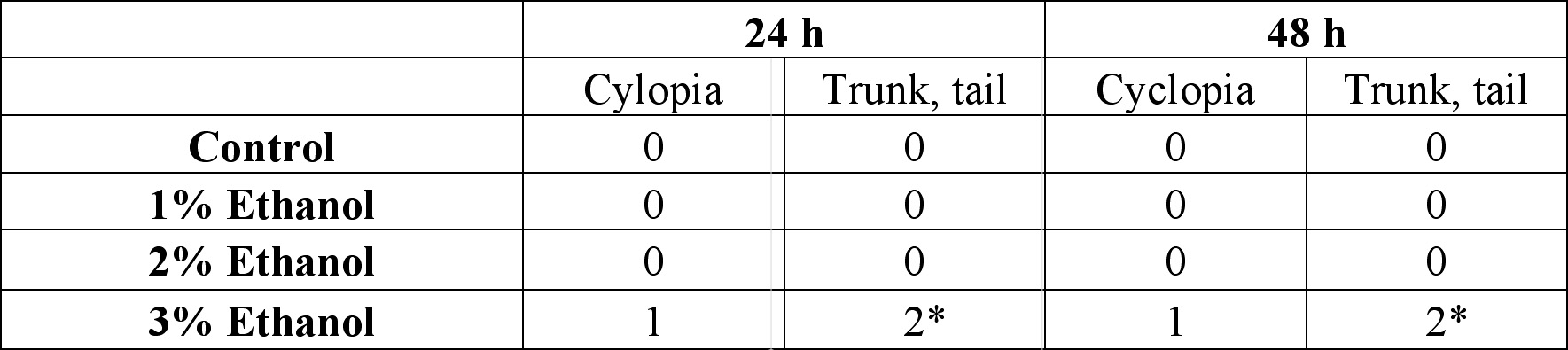
Physical deformities were observed only among the zebrafish embryos cultured in 3% ethanol-ZEM solution and not in 1% and 2%, suggesting that the critical ethanol concentration may be anywhere from 2% to 3%, including 3% (Table 1). That range is overlaps significantly with Blader and Strahle’s critical ethanol concentration of 2.4% (1998). However, the frequency of the deformities obtained in the experiment was different from Blader and Strahle’s 74.1% (1998). 48 hours after the ethanol treatment, at 3% ethanol solution, approximately 20% displayed any physical deformities. Such discrepancy may be due to mistakes made when staging the embryos for the experiment. According to Blader and Strahle, only ethanol treatment during a narrow time window comprising late blastula and early gastrula stages (dome/30% epiboly) causes cyclopia (1998). Moreover, deformities of the trunk and the tail also represent dome/30% epiboly stage treatment effects. Thus, it can be suggested that the embryos used in the experiment were generally past their 30% epiboly stages. However, the low percentage of deformed embryos is statistically insignificant due to the small sample size used in the experiment; only 10 embryos were used for each ethanol concentration as opposed to Blader and Strahle’s 700 (1998).
Though in low numbers, cyclopia was observed in embryos treated with 3% ethanol, supporting the fact that ethanol does induce cyclopia in zebrafish embryos (Blader and Strähle, 1998). Cyclopia is induced in amphibians and chick embryos when the prechordal plate is removed, suggesting that the prechordal mesoderm is required to separate the monolithic eye field into two lateral domains (Blader and Strähle, 1998). Therefore, it can be suggested that the embryo with separate, but more closely spaced eyes had its prechordal plate mildly affected by ethanol. The role of the prechordal plate is further evidenced in the cylopic embryo; it was clearly observed that what looked like a single medial eye was actually two eye vesicles fused at the center of the head when viewed under the microscope (Figures 1D, 2D). However, it is not clear as to why embryos treated with ethanol displayed significant lack of pigmentation when observed 24 hours after the ethanol treatment but not after 48 hours. One possible explanation could be that ethanol slowed down the zebrafish’s development. In the presence of ethanol, eye development may proceed abnormally slowly, resulting in retarding of the proliferation of the pigment cells of the eyes. Another possible explanation is that ethanol might have caused apoptosis of the eye pigment cells at 24 h and neighboring cells compensated for the loss by differentiating into eye pigment cells by 48 h.
Ethanol did cause defective development in the posterior structures in zebrafish embryos. Although the mechanisms behind the observed deformities of the trunk and the tail are not known, it can be conjectured that the impairment of epiboly may be responsible. Blader and Strahle noted that epiboly was marginally impaired by the treatment, with the advance of the blastoderm margin lagging behind untreated controls by 10 to 15% toward the end of gastrulation (1998). Epiboly establishes the posterior area of the embryo including the trunk region and the tail bud and, therefore, it seems reasonable that impairing the last 10-15% of epiboly may cause defective trunk and tail (Gilbert, 2003). In addition to a defective trunk and tail, it was also observed that there lack of pigmentation in the truck regions in embryos exposed to 3% ethanol (Figures 2D, 2E, and 4D). One possible explanation for the pigmentation deficiency is that ethanol might have caused deformities in the spinal cord from which neural crest cells destined to become melanocytes arise (Gilbert, 2003). However, it is not conclusive as to whether lack of pigmentation can be attributed to ethanol exposure. Largely due to small sample size, further experimentation is needed to confirm ethanol’s role in pigmentation defect.
Another noticeable effect of ethanol was the mortality rate (Table 2). Though Blader and Strahle specifically noted that ethanol does not seem to cause death in embryos, the experiment suggests that increasing ethanol concentration increased the number of deaths (1998). Six out of 10 embryos incubated in the 3% ethanol solution died as opposed to 1 out of 10 of the controls dying seem significant. Moreover, it is difficult to simply attribute the linear relationship displayed in the data to chance. It was conjectured that abnormal apoptosis, an accompanying symptom caused by ethanol exposure, may have caused the deaths of embryos. It is thought that the enzymes that cells use to degrade and clear ethanol out of the system produce free radicals which chemically react with important components of the cell, such as proteins, DNA, and lipids (Sulik et al., 1988). Therefore, it does not seem farfetched to suggest that ethanol induced apoptosis may ultimately cause death by damaging the essential cellular structures of the embryos.
This experiment demonstrated that ethanol induces physical deformities, more specifically cyclopia and trunk and tail deformities, in zebrafish embryos. It is known that, in humans, severe fetal alcohol syndrome causes mild forms of holoprosencephalies, a group of disorders that include cyclopia (Sulik et al., 1988). Therefore, the findings of this experiment seem to bear at least some relevance to experiments investigating fetal alcohol syndrome in humans. This experiment, however, did little to investigate the presence of ethanol induced abnormal apoptosis and its role on physical defects and deaths. In another experiment, embryos may be exposed to varying concentrations of ethanol, stained for apoptosis using vital stains such as the Acridine Orange, and observe whether there are correlation between deformities and cell death.
