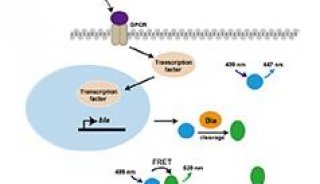
常见信号通路介绍
1. NF-κB信号
NF-kB(nuclear factor-kappa B)是1986年从B淋巴细胞的细胞核抽提物中找到的转录因子,它能与免疫球蛋白kappa轻链基因的增强子B序列GGGACTTTCC特异性结合,促进κ轻链基因表达,故而得名。它是真核细胞转录因子Rel家族成员之一,广泛存在于各种哺乳动物细胞中。迄今为止,在哺乳动物细胞内共发现5种NF-kB/Rel家族成员,它们分别是RelA(即p65)、RelB、C-Rel、p50/NF-kB1(即p50/RelA)和p52/NF-kB2。这些成员均有一个约300个氨基酸的Rel同源结构域(Rel homology domain, RHD)。这个高度保守的结构域介导Rel蛋白形成同源或异源二聚体,该结构域也是NF-kB与靶基因DNA序列的特异性结合区域。 细胞内NF-kB的活化过程受到精细调控。通常情况下,在细胞质中的NF-kB处于失活状态,与抑制蛋白IkB(inhibitory protein of NF-kB)结合成三聚体复合物。当出现TNF-a信号、炎症因子以及LPS、紫外线等外界刺激时,细胞因子与细胞膜表面的TNF受体结合后,TNF受体发生多聚化并与细胞质中TRADD分子发生相互作用。TRADD招募TRAF(TNFR-associated factor)和激酶RIP(receptor interacting protein),由RIP将信号传递给IKK(IkB kinase)。在NF-kB信号通路中IKK扮演了非常重要的角色,尽管上游信号路径的不同,但是最终都汇集到IKK。IKK由a、b和g三个亚基组成,作为激酶的IKK能使IkB的a亚基的Ser32和Ser36残基和b亚基的Ser19和Ser23残基磷酸化。IkB随即从p50/p65/IkB异源三聚体中解离出来,经泛素化修饰后通过蛋白酶体降解。于是,受到IkB抑制的NF-kB得以暴露其核定位序列(nuclear localization signals, NLS),迅速从细胞质进入细胞核内,与核内DNA上的特异序列相结合,从而启动或增强相关基因的转录。
2.JAK-STAT信号通路
1) JAK与STAT蛋白
JAK-STAT信号通路是近年来发现的一条由细胞因子刺激的信号转导通路,参与细胞的增殖、分化、凋亡以及免疫调节等许多重要的生物学过程。与其它信号通路相比,这条信号通路的传递过程相对简单,它主要由三个成分组成,即酪氨酸激酶相关受体、酪氨酸激酶JAK和转录因子STAT。
2) JAK-STAT信号通路
与其它信号通路相比,JAK-STAT信号通路的传递过程相对简单。信号传递过程如下:细胞因子与相应的受体结合后引起受体分子的二聚化,这使得与受体偶联的JAK激酶相互接近并通过交互的酪氨酸磷酸化作用而活化。JAK激活后催化受体上的酪氨酸残基发生磷酸化修饰,继而这些磷酸化的酪氨酸位点与周围的氨基酸序列形成"停泊位点"(docking site),同时含有SH2结构域的STAT蛋白被招募到这个"停泊位点"。最后,激酶JAK催化结合在受体上的STAT蛋白发生磷酸化修饰,活化的STAT蛋白以二聚体的形式进入细胞核内与靶基因结合,调控基因的转录。值得一提的是,一种JAK激酶可以参与多种细胞因子的信号转导过程,一种细胞因子的信号通路也可以激活多个JAK激酶,但细胞因子对激活的STAT分子却具有一定的选择性。例如IL-4激活STAT6,而IL-12却特异性激活STAT4。
3.Ras、PI(3)K和mTOR信号
随着人类基因组测序的完成,已发现了几百种蛋白激酶。根据它们结构上的相似性,这些激酶可分为多个蛋白家族,在细胞的增殖、生长、分化和凋亡等过程中发挥重要的生物学功能。Ras、PI(3)K和mTOR就是一类与细胞增殖紧密相关的蛋白激酶。真核细胞的正常生长受到周围环境所提供的养分的限制。Ras和PI(3)K信号通过调控下游分子mTOR,在调控细胞生长方面起着关键作用。在绝大多数的人肿瘤细胞中,Ras和PI(3)K信号通路中的关键调控因子都发生了明显的突变。究其原因,人们发现这条信号通路如果发生突变,就会导致细胞的存活和生长不再受到养分等环境条件的限制,进而诱导细胞癌变。值得注意的是,一些肿瘤抑制因子,如TSC1、TSC2和LKB1在营养匮乏的条件下减弱了mTOR信号通路的强度。相应地,TSC1、TSC2或者LKB1的失活突变,就会导致相似的癌症症状,并具有共同的临床表现。因此,这条确保细胞在环境适宜条件下发生增殖的信号通路,在被癌细胞利用后就可以使癌细胞在养料匮乏的条件下存活并生长。在筛选激酶抑制剂的过程中,人们设计了一系列针对mTOR、PI(3)K、RTKs和Raf等激酶的药物。在癌症的分子机理研究中,尽管这条信号通路研究得最透彻,但这些激酶在细胞和生物体内的生理功能远比我们想象的要复杂。
4.Wnt信号
Wnt信号通路广泛存在于无脊椎动物和脊椎动物中,是一类在物种进化过程中高度保守的信号通路。Wnt信号在动物胚胎的早期发育、器官形成、组织再生和其它生理过程中,具有至关重要的作用。如果这条信号通路中的关键蛋白发生突变,导致信号异常活化,就可能诱导癌症的发生。1982年,R. Nusse和H.E. Varmus在小鼠乳腺癌细胞中克隆得到第一个Wnt基因,最初它被命名为Int1(integration 1)。后来的研究发现小鼠Int基因与果蝇的无翅基因wg(wingless)为同源基因,因而将两者合称为Wnt。H.E. Varmus 本人也因他在癌症研究中的杰出贡献而获得1989年的诺贝尔生理医学奖。Wnt是一类分泌型糖蛋白,通过自分泌或旁分泌发挥作用。Wnt信号通路的主要成分包括:分泌蛋白Wnt家族、跨膜受体Frizzled家族、CK1、Deshevelled、GSK3、APC、Axin、β-Catenin、以及转录因子TCF/LEF家族。Wnt信号通路是一个复杂的调控网络,目前认为它包括三个分支:经典Wnt信号通路,通过β-Catenin激活基因转录;Wnt/PCP通路(planner cell polarity pathway),通过小G蛋白激活JNK(c-Jun N-terminal kinase)来调控细胞骨架重排;Wnt/Ca2+通路,通过释放胞内Ca2+来影响细胞粘连和相关基因表达。 一般提到Wnt信号通路主要指的是由β-Catenin介导的经典Wnt信号通路。
5. BMP信号通路
BMP(bone morphogenetic protein,骨形态发生蛋白)是TGF-β(转化生长因子,transforming growth factor-β)超家族中的重要成员。通过调节一系列下游基因的活性,控制着诸如中胚层形成、神经系统分化、牙齿和骨骼发育以及癌症发生等许多重要的生物学过程。BMP信号的传递主要通过配体BMP与细胞膜上的丝氨酸/苏氨酸激酶受体(BMP receptor, BMPR)特异性结合,形成配体受体二元复合物。同时,Ⅱ型受体(BMPR2)能够活化I型受体(BMPR1),并进一步将信号传递给细胞内的Smad分子。在BMP和TGF-β信号由细胞膜传递至细胞核的过程中,Smad蛋白起到了关键性的作用。活化的I型受体(BMPR1)进一步磷酸化Smad蛋白(Smad1、Smad5和Smad8),促使Smad分子从细胞膜受体上脱离下来,并在胞质内结合Smad4分子(common Smad,Co-Smad)后进入细胞核。在细胞核内,Smad多元复合物在其它DNA结合蛋白的参与下作用于特异的靶基因,调控靶基因的转录。
6. Ras2MAPK信号转导途径
1) Ras上游通路
Ras能被复杂的网络激活。首先,被磷酸化激活的受体,如PDGFR,EGFR直接结合生长因子受体结合蛋白(Grb2), 这些受体也可以间接结合并磷酸化含有src同源区2(SH2) 结构域的蛋白质(例如Shc,Syp)后,再激活Grb2.另外,Grb2的src同源区3(SH3)结构域与靶蛋白如mSos1, mSos2,C3G及发动蛋白(dynamin)结合.C3G与连接蛋白 Crk的SH3结构域结合后耦联酪氨酸磷酸化而激活Ras. Crk也能结合mSos1激活Ras.Grb2与激活的受体结合促 进鸟苷酸交换因子(Sos)蛋白定位在与Ras相邻的细胞膜上.这样,Sos与Ras形成复合体,GTP取代GDP与Ras结合后,Ras被激活,当GTP水解成GDP后Ras失活.Ras具有内在GTPase活性,它的活性可被RasGAPs调节,因而 RasGAPs扮演Ras活性调节剂的角色.另外,Ras失活也受 到高度调节.有三种蛋白质能水解GTP使Ras失活, 它们分别是P120GAP, neurofibromin和GAP1m,统称为RasGAPs.
2) Ras下游通路
Ras/Raf通路是最明确的信号转导通路.当GTP取代GDP与Ras结合,Ras被激活后, 再激活丝苏氨酸激酶级联放大效应,招集细胞浆内Raf1丝 苏氨酸激酶至细胞膜上,Raf激酶磷酸化MAPK激酶 (MAPKK), MAPKK激活MAPK. MAPK被激活后,转至细胞核内,直接激活转录因子.另外,MAPK刺激Fos,Jun转录因子形成转录因子AP1,该因子与myc基因旁的特异 的DNA序列结合,从而启动转录.myc基因产物也是转录 因子,它能激活其他基因.最终,这些信号集中起来诱导D 型Cyclin的表达和活性.D型Cyclin与Cyclin依赖性激酶 (如CDK4和CDK6)形成复合体,该复合体的形成促使细胞 从G1期进入S期.因此,Ras/Raf通路在受体信号和G1期 进展之间起着关键作用,然而, Ras/Raf通路不是调控G1 期进展的惟一通路.Ras与Raf单独结合不能促进Raf激酶活性,同时,Raf能被不依赖Ras的机制所激活(例如能被Src酪氨酸激酶和PKC所激活),MAPK也能被不依赖Ras机制(如通过调节整合素的活性)所激活.表明级联反应每一个信号蛋白质都能被多个上游蛋白质所激活,而它们也可能有另外的靶蛋白。