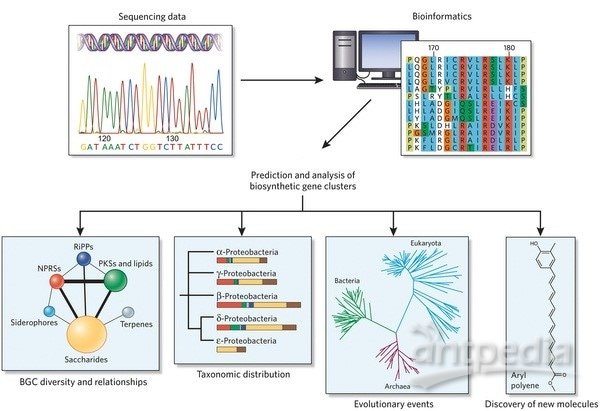
转化生长因子-β信号通路调控方式介绍
TGF-β信号通路参与许多细胞过程,因此受到频繁的调控。TGF-β信号通路有多种正反馈和负反馈调节机制,如配体和R-SMAD的激动剂,诱饵受体,R-SMAD和受体被泛素化等。
配体激动剂/拮抗剂
脊索蛋白和头蛋白都是骨形成蛋白(BMP)的拮抗剂。它们与BMP结合,阻碍其与受体的结合。有研究显示,脊索蛋白和头蛋白使中胚层形成背部组织/器官,二者都在非洲爪蟾的背唇中被发现,另外还将特定的上皮组织转化为神经组织。头蛋白在软骨形成和骨骼形成中起关键作用。
DAN家族的蛋白也与TGF-β家族成员拮抗。这些蛋白包括Cerberus,DAN以及Gremlin。这些蛋白包含可以形成二硫键的九个保守半胱氨酸。一般认为DAN与GDF5,GDF6及GDF7拮抗。
卵泡抑素与激活素结合并抑制激活素,它直接影响促卵泡激素的分泌。卵泡抑素也与前列腺癌密切相关,其基因的突变可能阻碍它对激活素的作用,而这种作用具有抗增殖的性质。
Lefty蛋白是TGF-β的一种调控因子,它参与了胚胎形成中的轴形成。它也是TGF超家族的成员。在鼠类胚胎中,这种蛋白在左侧不对称地表达,因而与左右分化相关。Lefty通过抑制R-SMAD的磷酸化产生作用,这样的过程通过一种持续性活化的TGF-βI型受体及Lefty活化的下游过程实现。
也有研究发现基于药物的拮抗剂,例如SB431542,它选择性地抑制ALK4, ALK5, and ALK7。
受体调控
转化生长因子受体3(TGFBR3)是发现的数量最多的TGF-β受体,它没有已知的信号域,但它能通过结合TGF-β并将其呈递给TGFBR2来增强TGF-β配体与TGF-βII型受体的结合。TGF-β信号通路的下游靶标之一GIPC与TGFBR3的PDZ结构域结合,抑制其蛋白酶体降解,进而增强TGF-β活性。它可能也是激活素RIL的抑制素复合受体。
BAMBI(BMP and activin membrane bound inhibitor,BMP和激活素膜结合抑制剂)与I型受体有相似的胞外域,它缺少一个丝氨酸/苏氨酸蛋白激酶胞内域,因此它是一个虚拟受体。BAMBI与I型受体结合,阻止其被激活。它是TGF-β信号通路中的负调控因子,可能在胚胎形成中抑制TGF-β表达。它的表达需要BMP信号。
FKBP12与I型受体的GS域结合,抑制II型受体将I型受体磷酸化。一般认为FKBP12及其同系物在没有配体时抑制I型受体活性,因为配体结合后它会断开与I型受体的连接。
R-SMAD调控
抑制性SMAD的功能
SMAD家族中还有两种抑制性蛋白(I-SMAD),即SMAD6和SMAD7。它们参与负反馈调节,并在TGF-β信号通路的调控中起关键作用。与其它SMAD蛋白一样,它们有一个MH1结构域和一个MH2结构域。SMAD7与其它R-SMAD蛋白竞争,与I型受体结合,并抑制其磷酸化。它们储存在细胞核中,一旦TGF-β被激活就进入细胞质并结合I型受体。SMAD6与SMAD4结合,阻碍其它R-SMAD蛋白与coSMAD蛋白结合。在TGF-β信号通路中,I-SMAD的表达水平提高,这意味着它是TGF-β信号通路的下游靶标。
R-SMAD的泛素化
E3泛素蛋白连接酶SMURF1和SMURF2调控SMAD的表达水平。与酶结合的E在酶上将泛素传递给RSMAD,导致其泛素化及蛋白酶体降解。SMURF1与SMAD1及SMAD5结合,而SMURF2 与SMAD1, SMAD2, SMAD3, SMAD6及SMAD7结合。SMURF2增强了SMAD7的抑制活动,而减少了SMAD2的转录活动。