题目:
Optically-controlled bacterial metabolite for cancer therapy. Nature Communications IF 12.124 iTRAQ
研究背景
随着对共生微生物认识的日益加深,人们逐渐意识到肿瘤内部是细菌和细胞共生的复杂微环境,并且相当一部分的细菌可以特异性地从循环系统中富集到肿瘤部位。在先前的研究中,科研人员也发现这些共生细菌在肿瘤的发展过程中起到关键性作用。在人为改造下,这一类共生细菌能够富集到肿瘤部位,抑制肿瘤生长,从而为肿瘤治疗提供全新方案。
近来,许多纳米级光催化材料如CdS和C3N4因其光电转换能力引起了广泛的关注,其可以将光能不断转换成电能。这些材料不仅能够增加细菌的代谢活性,而且能够避开负载极限,实现抗癌药物的富产。受上述光诱导还原的启发,本文作者将光催化系统与肿瘤靶向细菌融合,从而获得用于光控制的一氧化氮(NO)生成的生物/非生物混合物。首先合成了抑制自由基生成能力的碳量子点掺杂的碳化氮(CCN),以实现原位光电转换。然后,通过静电相互作用组装CCN和大肠杆菌(E. coli)以获得CCN@E. coli。本文中,作者提出了一个利用修饰的CCN@E. coli的光控细菌代谢物疗法(PMT)的概念。该技术利用修饰的CCN@E. coli将NO3-转化为在光照下具有癌症治疗效果的抗肿瘤一氧化氮(NO)。作者还利用iTRAQ定量蛋白质组学技术来研究其详细机制。PMT疗法优化了生物/非生物混合系统在生物医学中应用,转变了细菌癌症治疗方式。
研究内容及结果
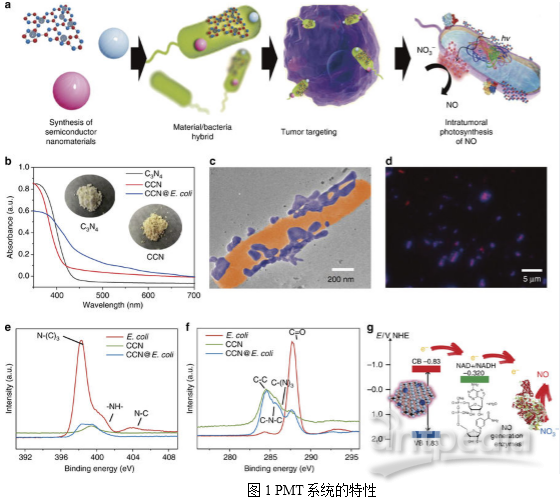
1. PMT系统的特性和机理
CCN@E. coli的制备如图1a所示。CCN通过静电相互作用与E. coli组装到一起。TEM图像显示修饰后,CCNs位于E. coli表面,在此过程中E. coli的形态没有发生变化(图1c)。如图1d所示,CCN和E. coli之间的空间重叠系数(0.94)也表明CCN@E. coli组装成功。作者推测,由于其合适的带隙,从CCN激发的光电子能够通过电子载体(例如NADH)转移至大肠杆菌的NO合成酶中。随后,NO3-可能以NADH依赖性方式酶促还原为NO(图1g)。因此作者利用经典的Griess方法研究了CCN@E. coli光学控制的NO释放行为(图2a)。
在正常生理条件下,L-精氨酸通过一氧化氮合成酶阳性细胞如巨噬细胞转化为NO。然而,由于来源有限,并且NO可能最终被氧化成无毒的NO3-,所以生理条件下产生的NO的抗癌潜能力很微弱。在CCN@E.coli辅助下的PMT疗法可将不可逆的肿瘤内NO代谢转化为循环反应,最大限度地提高NO的生物利用度。PMT系统的机制是一个两步过程:首先,E.coli利用发光的CCN光电产生的电子进行内源性NO3 -还原和NO生成(图2g),然后,由产生的NO引发细胞凋亡。
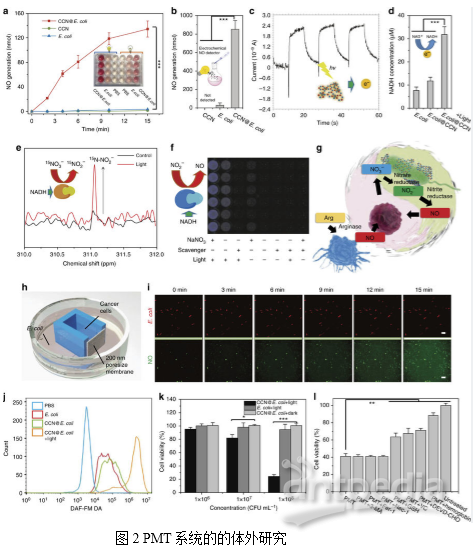
2. PMT的体外抗癌研究
为了使癌细胞和细菌在不直接接触的情况下实现代谢物的交换,作者使用3D打印技术制作了一种共培养装置。在这种培养体系中,NO可以通过CCN@E. coli(装置外培养)的多孔膜扩散到癌细胞(装置内培养),而CCN@E. coli或癌细胞都不能通过这个膜迁移。然后利用二氨基荧光素-FM二乙酸酯(DAF-FM DA)测定细胞内NO的浓度。如图2i所示,4T1细胞中绿色荧光的增加表明,PMT系统产生的NO能有效扩散到附近的腔室,上调癌细胞中的NO的水平。没有光照射的野生型大肠杆菌和CCN@E. coli不能使细胞内NO浓度增加(图2j)。
如图2k所示,使用CCN@E. coli光辐射的细胞存活率显著降低,在CCN@E. coli光辐射量为108 CFU/mL时,高达70%的4T1细胞能在24小时内被杀死。此外,作者还发现血红蛋白(一种NO清除剂)和Ac-DEVD-CHO(一种凋亡抑制剂)能有效地阻碍4T1细胞的凋亡(图2I)。基于上述观察,作者推断NO诱导的细胞凋亡是4T1细胞死亡的主要原因。
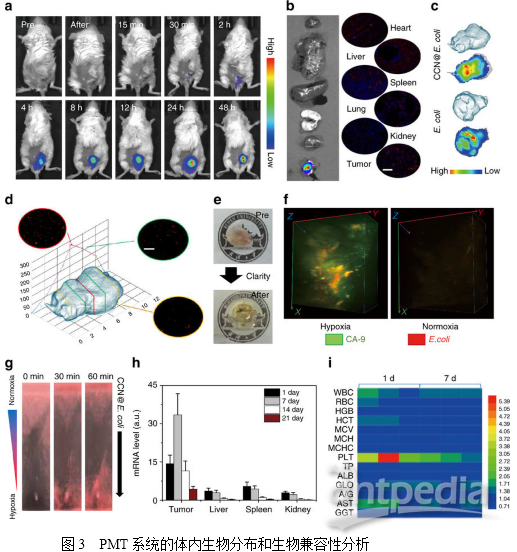
3. PMT系统的体内生物分布研究
为评估PMT系统的肿瘤靶向能力,将DIR标记的CCN@E.coli静脉注射到4T1荷瘤小鼠。如图3a所示,随着时间的延长,在肿瘤位置内检测到荧光的也逐渐增强。如图3b所示,离体荧光成像显示大量的细菌积聚在肿瘤位置,而肝脏或肾脏滞留的细菌几乎可忽略不计。值得注意的是,CCN的修饰并不影响E.coli的肿瘤靶向能力(图3c)。
通常,大多数合成药物载体由于其扩散限制,几乎不能穿透肿瘤组织。而作者推测PMT系统可能能够到达移植瘤的较深区域。结果表明,不同深度的肿瘤组织切片显示PMT系统分布于肿瘤组织中(图3d),并且发现移植瘤的中心聚积的细菌更多。为了全面揭示CCN@E.coli的肿瘤穿透性特征。作者使用表面活性剂辅助组织清除技术,即CLARITY,将肿瘤组织转化为透明的形式(图3e)。然后使用与肿瘤缺氧高度相关的生物标志物碳酸酐酶-Ⅸ(CA-9)对肿瘤缺氧区进行染色。从3D肿瘤荧光图像可观察到来自CCN@E.coli的红色荧光和来自CA-9的绿色荧光的共定位,而CCN@E.coli荧光可以在含氧量正常的区域发现(图3f)。进而证明了PMT系统可以充分利用大肠杆菌缺氧介导的趋化性,并且达到其他常规载体无法高效利用的缺氧区域。
随后,作者研究了CCN@E.coli在主要代谢器官如肝、脾和肾中的清除动力学。通过PCR和定性免疫荧光染色证明大部分细菌可以随着的时间延长而被清除。3周后,在免疫活性小鼠中可以检测到极少量的细菌(图3h),血液生化和血液学分析未发现PMT的长期副作用(图3i)。
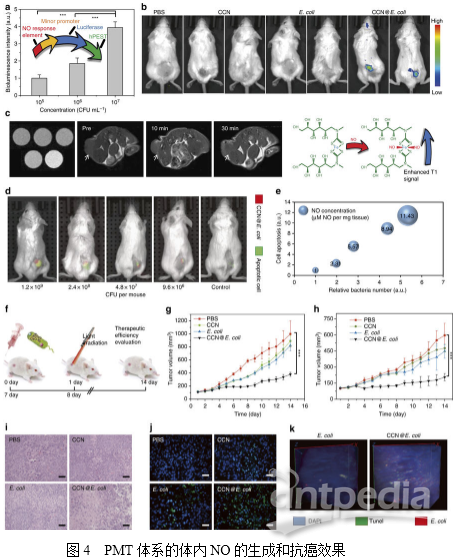
4. PMT系统中体内NO的生成
为了进一步阐明细胞光合行为,作者设计并构建了Nrf2受控萤光素酶表达质粒用以探测体内NO的产生,之后转染4T1细胞以构建4T1Nrf2细胞用于原位NO的检测。如图4a所示,4T1Nrf2-luc细胞暴露在CCN@E.coli环境下会导致其生物发光快速增加,作者还研究了4T1Nrf2-luc肿瘤小鼠中PMT系统的体内效率。如图4b所示,单独的CCN和E.coli都不能诱导产生可检测的NO。而CCN@E.coli处理组的生物发光强度显著增加。接下来,作者还合成了用于NO检测的磁共振成像(MRI)探针Fe-MGD。如图4c所示,作者发现在注射了CCN@E.coli和光照后,小鼠肿瘤内的T1信号增强,进一步证明了体内光控制的NO生成。
5. PMT的体内抗癌机制
作者利用缀合有Cy5.5的膜联蛋白V29来研究PMT的体内治疗反应。如图4d所示,随着CCN@E.coli注射剂量的增加,CCN@E.coli在肿瘤部位的累积增多。图4e的半定量分析结果也证明了肿瘤内细菌数目、肿瘤细胞毒性和NO浓度呈正相关。
此外,作者在4T1荷瘤小鼠和CT26荷瘤小鼠中检查了PMT的抗癌效率,治疗时间表如图4f所示。在4T1荷瘤小鼠中,PMT治疗抑制了79.3%的肿瘤生长(图4g)。而在CT26荷瘤小鼠中,PMT治疗抑制了70.2%的肿瘤生长(图4h)。这些结果表明PMT应该能够适用于治疗不同类型的癌症。
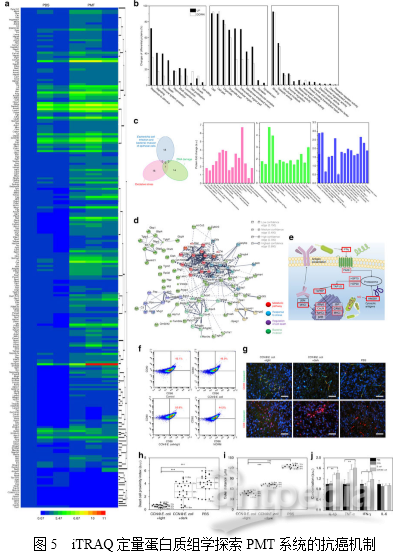
随后,作者利用定量蛋白质组学技术探索了PMT的详细抗癌机制。利用iTRAQ技术从4T1荷瘤小鼠癌组织(PMT处理组和PBS对照组)中共鉴定到4735个蛋白。如图5a所示,在PMT治疗后,肿瘤组织内鉴定到222个上调的差异蛋白和17个下调的差异蛋白(倍数变化≥1.5和P <0.05,超几何检验)。聚类分析和主成分分析结果显示PBS处理组和PMT处理组之间存在显著差异。对差异蛋白进行GO分析发现,刺激应答、信号、细胞死亡、免疫系统和细胞杀伤相关的蛋白质在PMT组中显著上调(图5b)。相反,与细胞增殖和生长相关的蛋白更多地发生下调。这些数据表明PMT在肿瘤中引起高应激反应,这主要是由于NO诱导的氧化损伤,并且与抗氧化活性相关的蛋白质也显著降低。通常,NO对癌细胞的细胞毒性主要归因于其诱导氧化应激和引发DNA损伤的能力。图5c所示的维恩图表明,大肠杆菌感染/侵入相关蛋白与DNA损伤/氧化应激相关蛋白之间没有重叠。因此,蛋白质组学研究表明CCN@E.coli对癌细胞的直接细胞毒性主要归因于NO本身。
有趣的是,在GO分析中发现了参与免疫应答的蛋白质水平显著升高。这一现象表明免疫系统可能有助于体内PMT的抗癌作用。PMT治疗后蛋白相互作用网络分析也可得出同样的结论。如图5e所示,KEGG分析发现抗原递呈途径被全面激活,并且证实了HMGB蛋白(一种参与触发抗原呈递的主导蛋白导致免疫原性细胞死亡)的上调。这些发现揭示了PMT可能通过HMGB引发MHC I类介导的途径诱导免疫原性细胞死亡。在体外模拟环境中,发现PMT治疗能够诱导DC突变(图5f)。这些结果表明,除了直接产生细胞毒性NO外,免疫应答也可能在在PMT的抗癌机制中起关键作用。
文章小结
一氧化氮(NO)在浓度较高的环境可以引发肿瘤细胞凋亡。为了实现对细菌合成NO能力的提升及控制,作者将碳量子点掺杂的碳化氮(CCN)负载到大肠杆菌(E.coli)MG1655上,利用合成材料良好的光催化性能,从而提高NO的产生,这一方法可以很好地实现细菌在肿瘤富集。本文作者全面研究了光控细菌代谢疗法(PMT)中NO的生成,细胞毒性细胞杀伤作用及其相关机制。动物实验证明,该疗法在小鼠肿瘤模型上表现出了约80%的抑瘤率。这一发现对哺乳动物-微生物的共生关系有了更深认识,并且极大丰富现有肿瘤疗法的内涵。定量蛋白质组学结果也表明免疫反应可能与该疗法相关, PMT疗法可能对癌症免疫疗法具有促进作用。该研究策略将为活体生物材料的设计和制备提供一种新的方法和视角。
参考文献
1. Treweek, J. B. et al. Whole-body tissue stabilization and selective extractions via tissue-hydrogel hybrids for high-resolution intact circuit mapping and phenotyping. Nat. Protoc. 10, 1860–1896 (2015).
2. Luo, C. H., Huang, C. T., Su, C. H. & Yeh, C. S. Bacteria-mediated hypoxiaspecific delivery of nanoparticles for tumors imaging and therapy. Nano Lett.16, 3493–3499 (2016).
3. Zheng, J. H. et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med. 9, eaak9537 (2016).
4. Zitvogel, L., Daillere, R., Roberti, M. P., Routy, B. & Kroemer, G. Anticancer effects of the microbiome and its products. Nat. Rev. Microbiol. 15, 465–478 (2017).
5. Wang, B. et al. Enhanced biological hydrogen production from Escherichia coli with surface precipitated cadmium sulfide nanoparticles. Adv. Energy Mater. 7, 1700611 (2017).
 引用
删除
18684909051 / 2018-08-21 08:08:01
引用
删除
18684909051 / 2018-08-21 08:08:01

