-
天然和人工培育冬虫夏草的比较蛋白质组学表征及其营养价值评估
2018-07-30 14:17:12
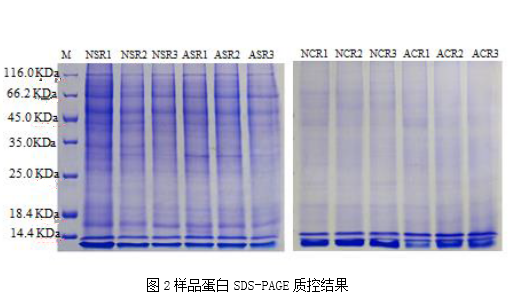
题目:A comparative proteomic characterization and nutritional assessment of naturally-and artificially-cultivated Cordyceps sinensis.Journal of Proteomics. IF 3.722 Label-free研究背景冬虫夏草因其营养和药用价值而受到越来越多的关注。近期,有报道称,成功通过人工方式培育出冬虫夏草。本文主要使用非标定量Label-free蛋白质组学技术研究自然生长和人工培育的冬虫夏草之间的蛋白质组学差异,揭示冬虫夏草蛋白质组学特征,了解其发育感染过程,并为人工培育提供参考信息。研究内容和结果1. 蛋白组学数据结果及蛋白质控人工培育的虫草和青藏高原上自然生长的虫草在形态特征高度相似(图1),并各自选择3个批次的样本进行蛋白提取。将虫草分为虫体和子座,分别用SDS-PAGE对样本蛋白进行质控检测(图2)。对解析的蛋白质组学数据进行质控分析,肽段质量容差为0.1Da,说明质谱鉴定肽段准确度高,肽段长度范围为8-38个氨基酸,分子量范围为400-12000Da,与预设分子量范围一致。在本次实验中,大多数鉴定蛋白含有2-20个unique肽段。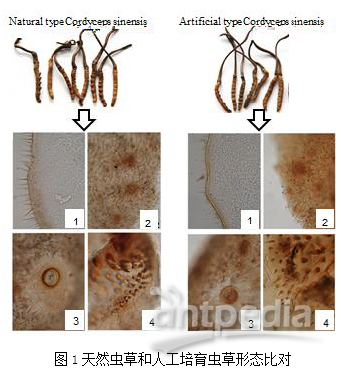
 使用变异系数(CV)验证实验方法的重复性,在本次实验结果中,人工培育的虫体(AC)、子座(AS)、天然生长的虫体(NC)、子座(NS)的CV值中位数分别为31.88%,27.44%,34.49%,34.94%。12个样品的相关性系数在0.68-0.93之间,这些结果证明了实验的可重复性。在所有的鉴定蛋白中,基于强度的绝对定量(iBAQ)值分布在1200中最丰富的蛋白质中(图3A)。所有样品和组内蛋白质的平均iBAQ值相似(图3B)。这些结果表明蛋白质组内比较可能不受蛋白质丰度系统差异的影响。
使用变异系数(CV)验证实验方法的重复性,在本次实验结果中,人工培育的虫体(AC)、子座(AS)、天然生长的虫体(NC)、子座(NS)的CV值中位数分别为31.88%,27.44%,34.49%,34.94%。12个样品的相关性系数在0.68-0.93之间,这些结果证明了实验的可重复性。在所有的鉴定蛋白中,基于强度的绝对定量(iBAQ)值分布在1200中最丰富的蛋白质中(图3A)。所有样品和组内蛋白质的平均iBAQ值相似(图3B)。这些结果表明蛋白质组内比较可能不受蛋白质丰度系统差异的影响。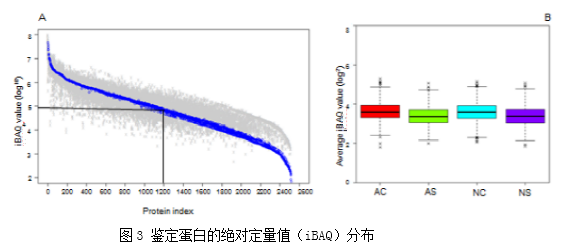 2. 天然虫草和人工培育虫草的蛋白表达图谱在本次实验中,12个样品共鉴定到22829条可信肽段(conf ≥ 95%),2541个蛋白质。使用PCA分析天然虫草和人工培育虫草蛋白表达图谱特征,发现虽然虫体和子座蛋白质分布有明显区别,但是天然虫草和人工培育虫草的整体蛋白质分布类似(图4)。COG功能注释到了1951个蛋白质。这些蛋白质主要参与能量产生、转化,氨基酸转运、代谢和转录调节(图5)。
2. 天然虫草和人工培育虫草的蛋白表达图谱在本次实验中,12个样品共鉴定到22829条可信肽段(conf ≥ 95%),2541个蛋白质。使用PCA分析天然虫草和人工培育虫草蛋白表达图谱特征,发现虽然虫体和子座蛋白质分布有明显区别,但是天然虫草和人工培育虫草的整体蛋白质分布类似(图4)。COG功能注释到了1951个蛋白质。这些蛋白质主要参与能量产生、转化,氨基酸转运、代谢和转录调节(图5)。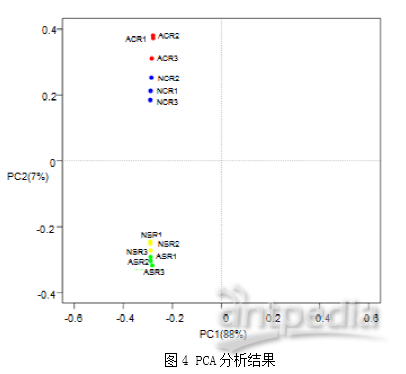
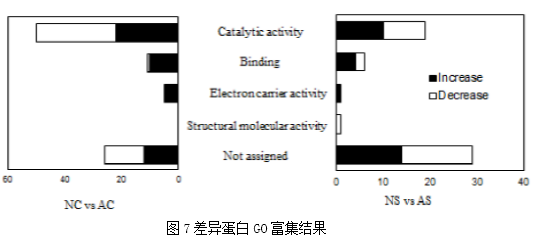
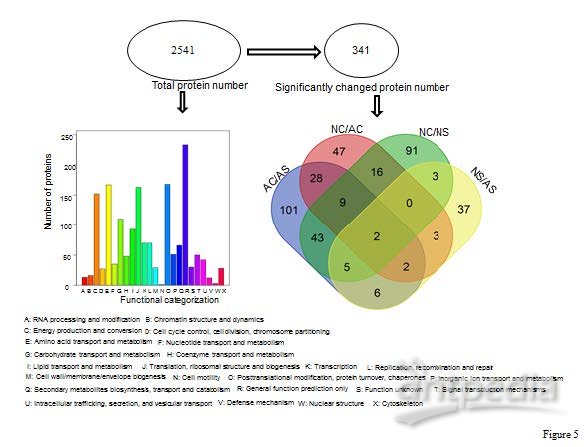 图5 COG功能注释结果3. 显著差异蛋白质鉴定结果本次实验中,显著差异蛋白数目分别为107(NC/AC)、58(NS/AS),195(AC/AS)、168(NC/NS)。使用层次聚类分析蛋白质表达变化趋势,主要聚成三类,Cluster I 由在AS中增加并在NC中减少的蛋白质组成;Cluster II包括在AC中降低并在NS中增加的蛋白质;Cluster III包括在AC或NC中增加并在AS和NS中均降低的蛋白质(图6)。为了确定这些蛋白质的功能特性,将它们的序列映射到GO数据库。GO富集表明这些差异丰富的蛋白质主要参与生物调节、单一生物过程,结合、催化活性,并定位于细胞器、细胞膜。结果说明这些显著差异蛋白在虫草生命活动中主要发挥催化和结合作用(图7)。
图5 COG功能注释结果3. 显著差异蛋白质鉴定结果本次实验中,显著差异蛋白数目分别为107(NC/AC)、58(NS/AS),195(AC/AS)、168(NC/NS)。使用层次聚类分析蛋白质表达变化趋势,主要聚成三类,Cluster I 由在AS中增加并在NC中减少的蛋白质组成;Cluster II包括在AC中降低并在NS中增加的蛋白质;Cluster III包括在AC或NC中增加并在AS和NS中均降低的蛋白质(图6)。为了确定这些蛋白质的功能特性,将它们的序列映射到GO数据库。GO富集表明这些差异丰富的蛋白质主要参与生物调节、单一生物过程,结合、催化活性,并定位于细胞器、细胞膜。结果说明这些显著差异蛋白在虫草生命活动中主要发挥催化和结合作用(图7)。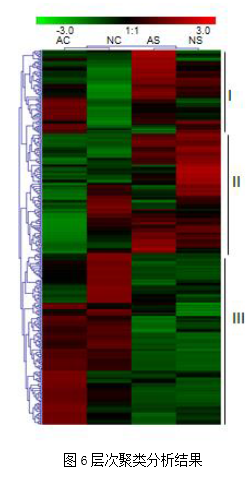
 4. 显著差异蛋白Pathway Enrichment 结果为了进一步研探索发生变化的生物学途径,使用KEGG Pathway 数据库进行富集分析,结果表明,天然虫草和人工培育虫草之间的差异蛋白主要参与次生代谢物合成、丙酮酸代谢、赖氨酸生物合成。在氨基酸合成、代谢途径中(图8),有7种相关蛋白发生表达量变化,其中,核糖5-磷酸异构酶(K01807)和丙酮酸激酶(K00873)在NC中降低,而NADH(P)结合蛋白(K00831)在NC中增加; NS中糖基转移酶(K00766)和雄性不育蛋白(K00143)增加;异柠檬酸/异丙基苹果酸脱氢酶(K00052)在NC中降低但在NS中升高,精氨酸琥珀酸合成酶(K01940)在NC和NS中均下降。
4. 显著差异蛋白Pathway Enrichment 结果为了进一步研探索发生变化的生物学途径,使用KEGG Pathway 数据库进行富集分析,结果表明,天然虫草和人工培育虫草之间的差异蛋白主要参与次生代谢物合成、丙酮酸代谢、赖氨酸生物合成。在氨基酸合成、代谢途径中(图8),有7种相关蛋白发生表达量变化,其中,核糖5-磷酸异构酶(K01807)和丙酮酸激酶(K00873)在NC中降低,而NADH(P)结合蛋白(K00831)在NC中增加; NS中糖基转移酶(K00766)和雄性不育蛋白(K00143)增加;异柠檬酸/异丙基苹果酸脱氢酶(K00052)在NC中降低但在NS中升高,精氨酸琥珀酸合成酶(K01940)在NC和NS中均下降。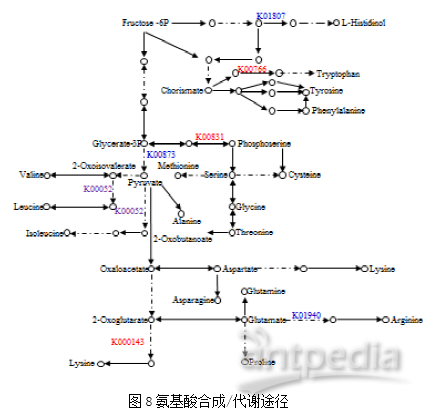 为了验证组学结果,LC-MS分析虫草代谢产物。根据代谢组学结果,共鉴定出8种氨基酸,其中4种氨基酸含量发生显著改变,具体为赖氨酸,精氨酸,苏氨酸和丝氨酸(图9)。氨基酸的变化趋势跟上游合成它们的酶的变化趋势是一直的。根据中国药典中关于测定核苷和核苷酸含量,特别是腺苷的规定,NC/NS以及AC/AS总离子色谱图中没有观察到显著差异(图10)。
为了验证组学结果,LC-MS分析虫草代谢产物。根据代谢组学结果,共鉴定出8种氨基酸,其中4种氨基酸含量发生显著改变,具体为赖氨酸,精氨酸,苏氨酸和丝氨酸(图9)。氨基酸的变化趋势跟上游合成它们的酶的变化趋势是一直的。根据中国药典中关于测定核苷和核苷酸含量,特别是腺苷的规定,NC/NS以及AC/AS总离子色谱图中没有观察到显著差异(图10)。
 虫草蛋白质中必须氨基酸比例与人类蛋白质中的比例相似,说明它具有营养价值。而且,天然虫草和人工培育虫草蛋白质中氨基酸成分和含量几乎相同,没有显著差异。文章小结该研究提供了天然虫草和人工培育虫草的蛋白表达图谱,发现天然虫草和人工培育虫草的真菌中的蛋白质和代谢产物组成相似,而虫体和子座的蛋白质和代谢产物方面则有区别。蛋白质组学结果表明氨基酸合成和代谢途径受到栽培方式的影响。 这些蛋白质组学数据将有助于理解冬虫夏草的药用价值,并为其人工培育提供参考。参考文献1. Shrestha UB, Bawa KS. Trade, harvest, and conservation of caterpillar fungus (Ophiocordyceps sinensis) in the Himalayas. Biol Conserv 2013;159:514.2. Zhao J, Xie J, Wang LY, Li SP. Advanced development in chemical analysis of Cordyceps. J Pharm Biomed Anal 2014;87:271–89.3. Hu HK, Xiao L, et al. Identification of chemical markers in Cordyceps sinensis by HPLC-MS/MS. Anal and Bioanal Chem 2015;407:8059–8066.4. Bereman MS, Beri J, Sharma V. An automated pipeline to monitor system performance in liquid chromatography-tandem mass spectrometry proteomic experiments. J Proteome Res 2016;15:4763-9.5. Xia E-H, Yang D-R, et al. The caterpillar fungus, Ophiocordyceps sinensis, genome provides insights into highland adaptation of fungal pathogenicity. Sci Rep 2017;7:1806.
虫草蛋白质中必须氨基酸比例与人类蛋白质中的比例相似,说明它具有营养价值。而且,天然虫草和人工培育虫草蛋白质中氨基酸成分和含量几乎相同,没有显著差异。文章小结该研究提供了天然虫草和人工培育虫草的蛋白表达图谱,发现天然虫草和人工培育虫草的真菌中的蛋白质和代谢产物组成相似,而虫体和子座的蛋白质和代谢产物方面则有区别。蛋白质组学结果表明氨基酸合成和代谢途径受到栽培方式的影响。 这些蛋白质组学数据将有助于理解冬虫夏草的药用价值,并为其人工培育提供参考。参考文献1. Shrestha UB, Bawa KS. Trade, harvest, and conservation of caterpillar fungus (Ophiocordyceps sinensis) in the Himalayas. Biol Conserv 2013;159:514.2. Zhao J, Xie J, Wang LY, Li SP. Advanced development in chemical analysis of Cordyceps. J Pharm Biomed Anal 2014;87:271–89.3. Hu HK, Xiao L, et al. Identification of chemical markers in Cordyceps sinensis by HPLC-MS/MS. Anal and Bioanal Chem 2015;407:8059–8066.4. Bereman MS, Beri J, Sharma V. An automated pipeline to monitor system performance in liquid chromatography-tandem mass spectrometry proteomic experiments. J Proteome Res 2016;15:4763-9.5. Xia E-H, Yang D-R, et al. The caterpillar fungus, Ophiocordyceps sinensis, genome provides insights into highland adaptation of fungal pathogenicity. Sci Rep 2017;7:1806. -
iTRAQ定量蛋白质组学:分析SEMA3B-AS1抑制人骨髓间充质干细胞成骨分化机制
2018-07-30 13:57:05
题目:SEMA3B-AS1-inhibited osteogenic differentiation of human mesenchymal stem cells revealed by quantitative proteomics analysis.Journal of cellular physiology. IF 3.923 iTRAQ研究背景常见的非编码RNA包括lncRNA、circRNA、miRNA,在细胞的生长、增殖、分化、凋亡中具有重要的调控作用。研究发现,非编码RNA与肿瘤的发生、发展、转移密切相关。作为非编码RNA的一员,lncRNA已被证实广泛在各类肿瘤的发生发展中扮演重要角色。新骨形成被认为是治疗骨相关疾病的一种新型的可替代方法,例如骨质疏松、骨架变形、骨折等。许多研究旨在提高新骨形成过程以希望治愈这些疾病。骨髓间充质干细胞(hMSC)是具有分化为成骨细胞、软骨细胞展现多种分化潜能的成纤维样多能干细胞,在新骨形成过程和骨质疏松发病等过程中发挥重要的作用。长链非编码RNA(lncRNAs)被发现参与成骨分化及骨质疏松的过程,比如H19通过吸附miRNA-675促进成骨细胞分化,MEG3通过靶向BMP4或者miR-140-5p来提高MSC的骨生成。但是lncRNA参与hMSC分化过程以及具体的机制目前不是很清楚。hMSC的分化是接受不同环境刺激后发生的过程,细胞表面受体在这个过程中发挥重要的作用,可以起到信号级联及放大的作用。SEMA家族蛋白被发现在骨生成或者骨疾病中发挥作用。如Sema6d、mema4d、sema3a等被发现可以平衡破骨细胞和成骨细胞来调节骨形成和骨吸收。前期研究发现SEMA3B-AS1可以抑制hMSC增殖和骨分化,具体机制仍需研究。目前lncRNA研究手段主要有:① 功能、表型、疾病性相关研究;②iTRAQ蛋白质组学分析lncRNA差异表达、预测新的lncRNA、生信分析lncRNA靶基因;③ 互作研究及质谱寻找及验证与lncRNA结合的RNA及蛋白等。肿瘤等疾病中lncRNA作用机制研究有望为疾病的治疗寻找新的靶点。本研究主要采用iTRAQ蛋白组学研究反义lncRNA-SEMA3B-AS1的作用机制。研究内容及结果1. MTT法检测细胞增殖和茜素红S染色实验证实SEMA3B-AS1可以抑制细胞的增殖(a)和向成骨转化(b)。说明SEMA3B-AS1具有抑制人骨髓间充质干细胞增殖和向成骨分化的功能作用。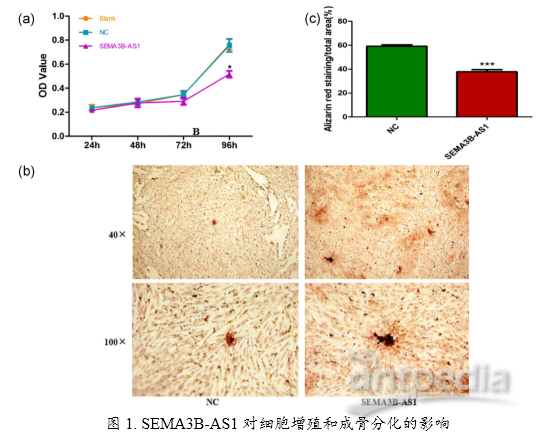 2. 上述功能验证实验已经确定了SEMA3B-AS1对人骨髓干细胞的增殖和分化产生了影响。但是SEMA3B-AS1通过何种途径发挥功能呢?作者接着包装SEMA3B-AS1过表达慢病毒,采用iTRAQ蛋白组学技术检测过表达SEMA3B-AS1和空载体的两个细胞组中差异蛋白变化。通过对差异蛋白做了聚类和功能富集,发现SEMA3B-AS1可能通过上调或下调相关基因来影响肌动蛋白骨架、黏着斑、胞外基质与受体的相互作用,来调节细胞的骨生成,进而影响细胞增殖和分化。
2. 上述功能验证实验已经确定了SEMA3B-AS1对人骨髓干细胞的增殖和分化产生了影响。但是SEMA3B-AS1通过何种途径发挥功能呢?作者接着包装SEMA3B-AS1过表达慢病毒,采用iTRAQ蛋白组学技术检测过表达SEMA3B-AS1和空载体的两个细胞组中差异蛋白变化。通过对差异蛋白做了聚类和功能富集,发现SEMA3B-AS1可能通过上调或下调相关基因来影响肌动蛋白骨架、黏着斑、胞外基质与受体的相互作用,来调节细胞的骨生成,进而影响细胞增殖和分化。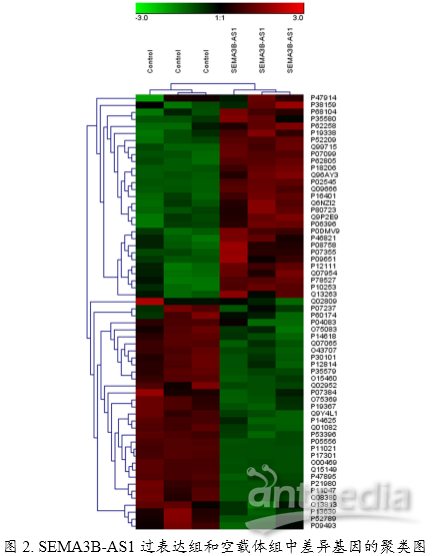 图3. 功能富集和差异蛋白表达分析
图3. 功能富集和差异蛋白表达分析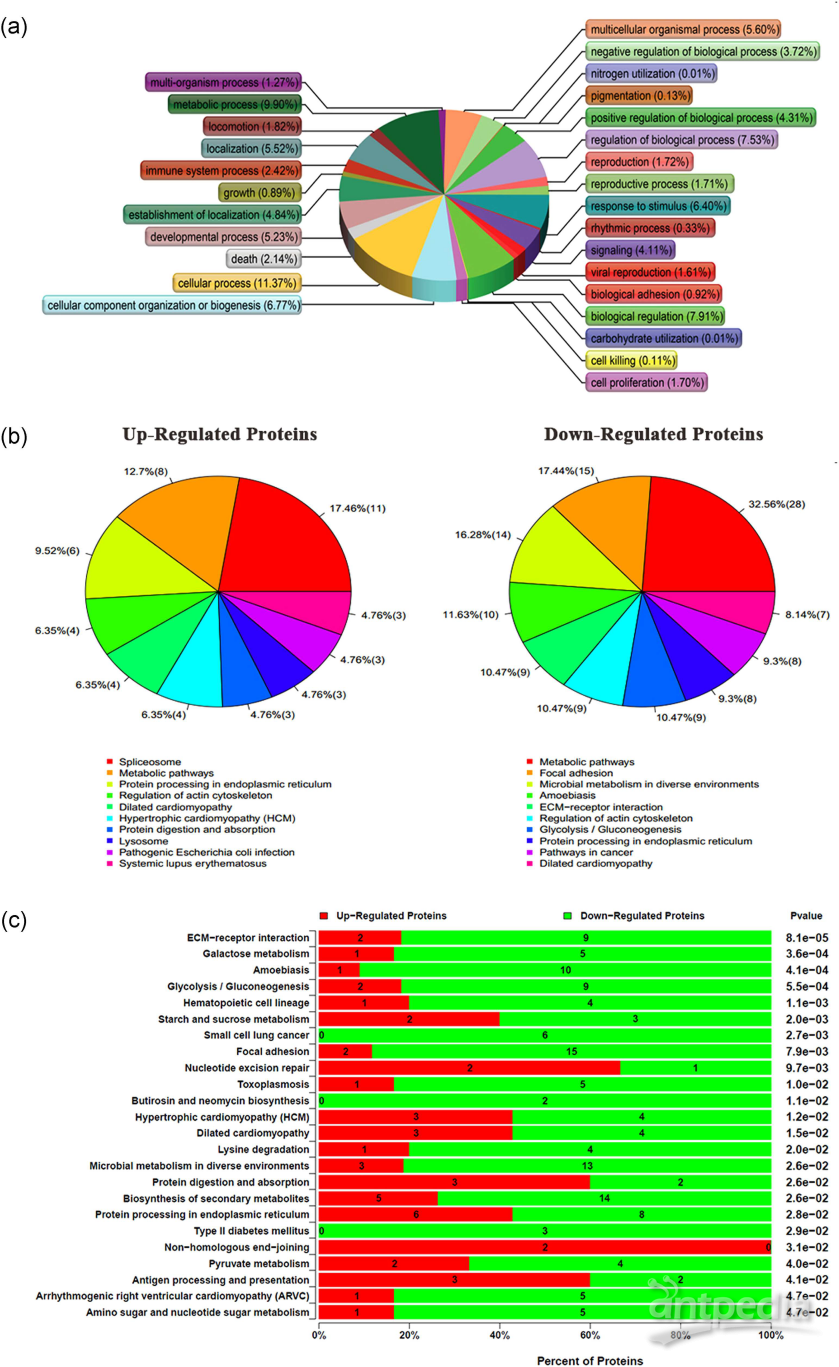 图3. 功能富集和差异蛋白表达分析文章小结lncRNA分为正义lncRNA和反义lncRNA,同正义lncRNA相比,反义lncRNA由于与正义链互补,反义链lncRNA与相邻或相近基因的调控关系更加密切。首先,作者以骨质疏松发病机制研究为目的,以人骨髓间充质干细胞、反义lncRNA(SEMA3B-AS1)为对象,进行功能实验,检测了SEMA3B-AS1对细胞增殖和骨生成的影响。同时,作者采用iTRAQ蛋白组学技术检测分析受SEMA3B-AS1调控的蛋白差异表达及差异蛋白的生物学过程及相关的代谢通路。最后,作者分析了SEMA3B-AS1调节的骨生成在骨质疏松发生中的作用,得出SEMA3B-AS1可能是骨质疏松治疗的一个靶点。参考文献1. J.C. Chen, C.R. Jacobs, Mechanically induced osteogenic lineage commitment of stem cells, Stem Cell Res Ther, 4 (2013) 107.2. J.J. Quinn, H.Y. Chang, Unique features of long non-coding RNA biogenesis and function, Nat Rev Genet, 17 (2016) 47-62.3. C. Sang, Y. Zhang, F. et al , Tumor necrosis factor alpha suppresses osteogenic differentiation of MSCs by inhibiting semaphorin 3B via Wnt/beta-catenin signaling in estrogen-deficiency induced osteoporosis, Bone, 84 (2016) 78-87.4. P.S. Mathieu, E.G. Loboa, Cytoskeletal and focal adhesion influences on mesenchymal stem cell shape, mechanical properties, and differentiation down osteogenic, adipogenic, and chondrogenic pathways, Tissue Eng Part B Rev, 18 (2012) 436-444.
图3. 功能富集和差异蛋白表达分析文章小结lncRNA分为正义lncRNA和反义lncRNA,同正义lncRNA相比,反义lncRNA由于与正义链互补,反义链lncRNA与相邻或相近基因的调控关系更加密切。首先,作者以骨质疏松发病机制研究为目的,以人骨髓间充质干细胞、反义lncRNA(SEMA3B-AS1)为对象,进行功能实验,检测了SEMA3B-AS1对细胞增殖和骨生成的影响。同时,作者采用iTRAQ蛋白组学技术检测分析受SEMA3B-AS1调控的蛋白差异表达及差异蛋白的生物学过程及相关的代谢通路。最后,作者分析了SEMA3B-AS1调节的骨生成在骨质疏松发生中的作用,得出SEMA3B-AS1可能是骨质疏松治疗的一个靶点。参考文献1. J.C. Chen, C.R. Jacobs, Mechanically induced osteogenic lineage commitment of stem cells, Stem Cell Res Ther, 4 (2013) 107.2. J.J. Quinn, H.Y. Chang, Unique features of long non-coding RNA biogenesis and function, Nat Rev Genet, 17 (2016) 47-62.3. C. Sang, Y. Zhang, F. et al , Tumor necrosis factor alpha suppresses osteogenic differentiation of MSCs by inhibiting semaphorin 3B via Wnt/beta-catenin signaling in estrogen-deficiency induced osteoporosis, Bone, 84 (2016) 78-87.4. P.S. Mathieu, E.G. Loboa, Cytoskeletal and focal adhesion influences on mesenchymal stem cell shape, mechanical properties, and differentiation down osteogenic, adipogenic, and chondrogenic pathways, Tissue Eng Part B Rev, 18 (2012) 436-444. -
蛋白质组学研究:揭示人调节T细胞在保护细胞信号通路中的适应性
2018-07-30 13:35:59
Proteomic Analyses of Human Regulatory T Cells Reveal Adaptations in Signaling Pathways that Protect Cellular Identity.Immunity IF19.734 蛋白质组研究背景
Treg cells(调节性T细胞)因其转录因子FOXP3的表达和抑制免疫应答的能力构成了独特的CD4+T细胞谱系。Treg 细胞可以保护免疫耐受,抑制免疫细胞造成的组织损伤,促进组织修复。Treg细胞也有不好的一面,它可以阻碍机体对癌细胞的免疫能力,有研究认为Treg这部分的功能缺失可能是造成人类自身免疫疾病和过敏发生的基础。因此,需要有针对性的促进或抑制人类疾病中的Treg细胞功能。Treg细胞和Tconv细胞密切相关,作者希望能够在分子水平上对二者进行详细研究和区分。研究内容及结果
1. Treg细胞蛋白表达特征
作者从健康人供体的外周血单核细胞(PBMCs)分离CD4 + T细胞亚群。这些细胞亚群包含nTconv细胞(CD45RA +CD25-)、记忆(m)Tconv细胞(CD45RA-CD25-)、nTreg细胞(CD45RA+CD25hi)和eTreg细胞(CD45RA-CD25hi)(图1A)。两种Treg细胞亚群都表达FOXP3和Helios并且缺乏IL-7受体α链(CD127),而nTconv和mTconv细胞亚群则相反,它们缺乏FOXP3和Helios,但是能表达CD127。使用高分辨率质谱(MS),作者鉴定了平均35,744±3,757个肽段,5,955±344个蛋白gurup。在所有五个CD4+T细胞亚群中定量了4,358种不同的蛋白质。其中,422种蛋白质基于其非标记定量(LFQ)值在CD4+T细胞亚群中表现出差异表达(FDR <0.05)。 总数据集的主成分分析(PCA,图1B)、Pearson相关分析(图1C)、差异表达蛋白的层次聚类(图1D)证实了生物重复的密切相关性。作者分离CD4+T细胞亚群的原则并不是基于细胞谱系,而是根据CD45RA的表达(图1D)。富集分析显示,幼稚Treg和Tconv细胞共享参与磷酸戊糖代谢、NADP代谢和染色质组织等过程中的过表达蛋白(图1D),而eTreg和mTconv细胞共享参与蛋白质合成、细胞运输、信号传导和凋亡的过表达蛋白(图1D)。基于分化阶段蛋白质表达的相似性可部分反映淋巴组织(幼稚细胞)与非淋巴组织(效应细胞)的优先定位。从聚类结果可以发现具体的Treg细胞蛋白特征,如nTreg和eTreg细胞共享某些高表达蛋白(cluster1)和低表达蛋白(cluster9-10),而eTreg细胞则独自表现出在cluster2中蛋白高表达以及在cluster6中蛋白低表达。 图1 CD4+T细胞亚群蛋白质组学结果2. Treg细胞的mRNA表达特征
图1 CD4+T细胞亚群蛋白质组学结果2. Treg细胞的mRNA表达特征
Treg细胞的蛋白组学结果与已发表的相似的细胞亚群的转录组学结果关联性较低。为了进行直接比较,作者对五个CD4+T细胞亚群进行了全基因组mRNA深度测序。也基于它们的转录组分析发现,nTreg和nTconv细胞聚集在一起并远离三种效应和记忆表型CD4+T细胞群(图2A)。在五个CD4 + T细胞亚群(图2B)之间总共649个差异表达mRNA(p <0.05),包括预期的标志基因如IL-7R、IKZF2(HELIOS)和效应细胞因子。作者分析发现,eTreg细胞的mRNA表达特征既有特异性(图2B,cluster7;),也有和其他Treg细胞的一致的共性,如都可以表达FOXP3、IL2RA、TIGIT等分子(图2B和2C,cluster6)。已经发表的文章结果也证明了作者的发现。作者发现,Treg细胞蛋白质组学结果和转录组结果中只有三个分子(FOXP3,SHMT2和SWAP70)表达量趋势上发生重叠(图2F)。事实上,553 个mRNA和409个蛋白质在五个CD4+ T细胞亚群之间显着差异表达,并且可以在两个水平上定量,但是二者仅重叠48个(图2G)。因此,与众多报道的文献结果一致,相对于转录组结果,蛋白质组学会获得明显不同的结果。 图2 CD4+ T细胞亚群mRNA表达结果3. 蛋白质水平与mRNa水平的区别
图2 CD4+ T细胞亚群mRNA表达结果3. 蛋白质水平与mRNa水平的区别
为了定量比较蛋白质和mRNA水平,作者使用基于强度的绝对定量分析蛋白质数据(iBAQ)。发现蛋白质和mRNA的丰度水平都超过五个数量级(图3A和3D)。在蛋白水平中,丰度最高的是核糖体和代谢蛋白(图3B),在mRNA水平中,丰度最高的是编码参与(免疫)细胞、信号传导和功能的核糖体组分和分子(图3E),差异蛋白的丰度通常要比mRNA要高。
作者对4792个mRNA-protein对的表达量在转录组和蛋白组水平上进行定位,发现其相关系数约为0.44±0.01。422种差异表达的蛋白质中,在mRNA水平检测到409种,并进行定量比较(图3G)。虽然蛋白质和mRNA表达水平通常相关(例如,FOXP3I、KZF2),但是大多数仅有一种(蛋白或mRNA)的表达量达到差异统计标准,这就解释了差异表达的蛋白质和mRNA之间的有限重叠(图2F)。作者发现一些分子仅在mRNA或蛋白质水平上真正差异表达(图3H),这说明细胞调节具有一定的层次范围。例如,对于FTH1(图3G),是一个已知在严格的翻译控制下的蛋白质(Hentze等人,2010),它在nTreg和eTreg细胞中丰度都很高;STAM2在mTconv细胞中虽然mRNA水平低,但是蛋白丰度很高(图3G), 另一方面,在nTreg和eTreg细胞中,mRNA上高表达但蛋白质水平并不高(图3G)。作者认为联合蛋白质组和转录组分析可以增加对通路分析的可信度。这样的组合数据有力地论证了与其他CD4+ T细胞亚群相比,核因子kB(NF-kB)和JAK-STAT途径在eTreg细胞中更容易脱敏(图3I)。总之,其研究结果强调了蛋白质组学分析对细胞类型功能表征的重要性。 图3 mRNA和蛋白表达的通路富集分析结果4. Treg细胞具有稳定的蛋白质特征
图3 mRNA和蛋白表达的通路富集分析结果4. Treg细胞具有稳定的蛋白质特征
作者定义的常见Treg细胞特征由22个具有较高表达的蛋白质和29个低表达蛋白质组成(图4A)。 免疫印迹和流式细胞术证实了这些蛋白质中的8种蛋白质组学数据。 在早期的Treg细胞蛋白质组学研究中,多是基于CD4+CD127-CD25+和CD4+CD127+ CD25-细胞,覆盖的蛋白质组较少,所以并没有找到类似作者发现的组学特征。常见的Treg细胞特征包括FOXP3、IKZF2(Helios)、代谢蛋白GK、UGP2和SHMT2、铁储存蛋白铁蛋白重链和轻链(FTH1,FLT),以及溶酶体蛋白ASAH1、GGH、GUSB、SGSH和PLBD2,与Tconv细胞相比,它们都在Treg中以高丰度表达(图4A和4B)。 该特征还包括糖酵解酶HK1、ME2、脂肪酸氧化酶AP00和线粒体脂肪酸转运蛋白(CPT1A),与Tconv细胞相比,均在Treg中以低丰度表达。作者发现许多信号分子在常见的Treg细胞特征中是差异表达的,如脱氢泛素酶OTULIN的高表达,TNFα诱导的NF-kB活化的抑制剂和TNFRSF1A接头TRADD的低表达(图4A和4B),以及mRNA水平的TNFRSF1B的高表达,表明Treg细胞在TNFR信号传导中表现出适应性。脂质磷酸酶INPP5D(SHIP-1)的高表达抑制PI3K-AKT信号传导,mTOR活化剂RPS6KA1和RPS6KA3的低表达,同样也表明PI3K / AKT / mTOR途径的适应性,Treg细胞中STAT4和NFATc2的低表达突出。为了确定上文鉴定的蛋白质表达模式的稳定性,作者在体外T细胞扩增后进行蛋白质组学分析。通过CD3、CD28和IL-2刺激将nTreg和nTconv细胞在体外扩增2周,孵育4天,并通过FOXP3、Helios、CTLA-4和CD25染色以及FOXP3基因TSDR甲基化验证它们的身份。值得注意的是,即使在通过CD3和CD28途径重新激活后,Treg细胞核心特征中大多数蛋白质特异性表达模式也基本上是保守的(图4C,4D)。作者认为Treg细胞在某些代谢功能以及重要信号通路的组成中与Tconv细胞本质上不同。 图4 常见的Treg细胞蛋白组学特征5. eTreg细胞特有的蛋白质组学特征
图4 常见的Treg细胞蛋白组学特征5. eTreg细胞特有的蛋白质组学特征
除了常见的Treg细胞特征外,作者发现eTreg细胞与其他CD4+ T细胞亚群相比有独特的蛋白质簇表达特征:具有相对高的(eTreghi)和低的(eTreglo)表达(图5A)。eTreghi簇包括参与DNA复制(MCM2,-3,-4,-6,-7,FEN1)、有丝分裂(CORO1C,TUBA1B,TUBB,MYH9)(图5A和5B)和细胞凋亡(FAS,CASP3)的蛋白质,这和已有研究一致,说明这些细胞正在发生分裂。在eTreghi簇中的41种蛋白质中,17种在体外Treg细胞扩增后仍显示出上升趋势,其中5种达到统计学显着性。同样,eTreglo簇中39种蛋白中的25种在体外扩增的Treg细胞中保持低表达。eTreglo簇包含多种调节细胞凋亡敏感性的GIMAP(图5A和5B)。重要的是,eTreglo簇中几乎40%的蛋白质在细胞信号传导中具有功能,它们包括NF-kB途径的多种成分(PRKCB、DPP4、NF-kB1、NF-kB2)、细胞因子受体途径(IL-7R、INPP4B、STAM2、STAT3)和TCR途径(TRAT1、VAV1、THEMIS)。即使在体外培养之后,Treg细胞中许多蛋白质的表达相对较低,表明这种对信号通路的明显脱敏作用,并不仅仅是通过这些通路在体内激活信号的负面反馈调节。
FOXP3可以与关键转录因子(如IL17A和IL4)发生物理相互作用并且具有可以淬灭它们反式激活Tconv细胞效应基因的能力。反之亦然,如YY1的因子可以抑制FOXP3介导的Treg细胞程序的控制。作者为了确定Treg细胞中FOXP3的相对浓度是否足以“压倒”这样的相互作用因子,使用蛋白质组学标尺方法确定了每个细胞的蛋白质拷贝数,发现FOXP3在eTreg细胞中的拷贝数超过其许多伴侣蛋白,其中差别最大的是转录因子,如NFATc1和STAT4(图5 C)。然而,对于一些转录因子(YY1、RUNX1和NFATc2),FOXP3过量仅为2至3倍,这可以解释为什么FOXP3的适度减少允许Treg细胞产生效应细胞因子。最后,作者发现mTconv细胞中FOXP3的表达水平可能不足以有效中和其伴侣转录因子(图5C)。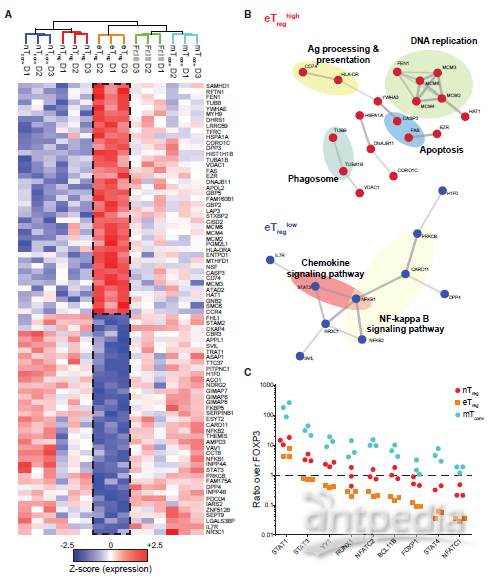 图5 eTreg细胞蛋白特征6. eTreg细胞具有效应基因表达的迟钝途径
图5 eTreg细胞蛋白特征6. eTreg细胞具有效应基因表达的迟钝途径
作者发现与Tconv细胞相比,eTreg细胞的IPA分析显示TCR、模式识别受体(PRR)、细胞因子受体和TNFRSF家族诱导的NF-κB途径信号传导具有适应性。此外,通过NF-kB1和NF的流式细胞术证实,eTreg细胞中NF-kB1(p50)、NF-kB2(p52)和RELA(p65)的mRNA水平较低。因此,在eTreg细胞中,NF-kB1的反cd3和反cd28诱发的核核易位被削弱了,但在nTreg细胞中却没有。NFATc1和NFATc2蛋白水平在nTreg细胞和eTreg细胞中都较低。此外,NFATc2在普通Treg细胞的蛋白中含量较低,在Treg细胞在体外(图4A和4D)后保持低表达。最后,在eTreg细胞转录组(图6A)中,带有NFATc2绑定元素的基因含量降低(图6A),这表明该因素在体内的这些细胞中不那么活跃。炎性细胞因子控制Treg细胞行为可能会对其稳定性造成影响,在传递细胞因子受体信号的STAT转录因子中,STAT3、STAT6,尤其是STAT4在eTreg细胞中表现出低表达。STAT4表达在mRNA和蛋白质水平均较低,STAT4的低蛋白质表达是常见Treg细胞蛋白特征的一部分。在eTreg细胞转录组中,带有STAT4结合元素的基因也没有得到充分的表达(图6A),这支持了体内eTreg细胞中STAT4活性降低的观点。通过响应IL-12和I型IFN磷酸化Y693,在人CD4 + T细胞中激活STAT4,后者在人CD4 + Tconv细胞中诱导最强的早期STAT4磷酸化。这些细胞因子一致地诱导Treg细胞中STAT4的磷酸化比从血液中新鲜分离的Tconv细胞中更少(图6C),并且这种差异在体外扩增的细胞中甚至更为显着(图6E)。STAT4是IFNg基因表达的主要调节因子,因此,作者推断STAT4的低表达可能使Treg细胞与炎性细胞因子诱导IFN-g隔离。与此假设一致,IFN-α并且IL-12不能在Treg细胞中诱导IFN-g产生(图6F)。然而,当故意过表达STAT4时,这些细胞因子确实激发了IFN-g的产生Treg细胞(图6G)。值得注意的是,尤其是当用IFN-α或IL-12刺激Treg细胞时,STAT4的过表达也诱导了IL-2的产生(图6G)和FOXP3表达的显着丧失(图6H和6I)。这些发现表明STAT4的低表达有助于在炎性环境中保护Treg细胞。
因为Treg细胞需要来自炎性细胞因子的输入,所以虽然STAT4表达低,但是作者仍验证了Treg细胞是否能够对I型IFN起反应。结果发现,在Treg和Tconv细胞中均等地诱导STAT1磷酸化(图6B和6D)。 此外,IFN-α容易在抗CD3和抗CD28活化的Treg细胞中诱导转录因子T-bet和趋化因子受体CXCR3的表达(图6J,6K)。 总之,这些发现表明STAT4的选择性低表达允许Treg细胞响应炎性细胞因子(例如归巢至发炎组织),而不损害Treg细胞。 图6 Treg细胞中选择性缺乏STAT4激活7. 在Fr.III细胞中可以区分不同的细胞群
图6 Treg细胞中选择性缺乏STAT4激活7. 在Fr.III细胞中可以区分不同的细胞群
Fr.III细胞是CD4+CD25+并且表达FOXP3,可以产生炎性细胞因子并且在体外缺乏强大抑制能力的细胞。该群体中的许多细胞表达CD127表明它是含有类似Treg或Tconv细胞的细胞的混合群体。为了更好地表征这一群体,作者根据CD127是否表达将其分成两个部分,然后进行每个级分的蛋白质组学分析,并与nTconv、mTconv、nTreg和eTreg细胞的蛋白质组一起进行分析。层次聚类分析和PCA显示CD127+ 亚群与mTconv细胞密切相关(图7A和7B),说明CD127用于区分Tconv和Treg细胞有用性。值得注意的是,CD127- 亚群与eTreg细胞几乎无法区分。CD127- Fr.III和eTreg细胞之间的紧密蛋白质组关系表明前者可能是真正的Treg细胞群,然而,FOXP3中的TSDR在CD127- Fr.III细胞中比在eTreg细胞中更加甲基化(图7C)。此外,CD127-Fr.III细胞含有大部分产生一种或多种效应细胞因子的细胞(图7F),这与eTreg细胞不同,因为eTreg细胞大多缺乏这种能力。并非CD127- Fr.III群体中的所有细胞都产生效应细胞因子,也并非eTreg细胞群中的所有细胞都缺乏产生此类细胞因子的能力,这表明即使这些明确定义的细胞群体仍可能是异质的。因此,作者在蛋白质组数据集中搜索了可能有助于区分具有不同功能特性的细胞的标记物。之前已有研究报道,在Treg细胞上发现的两种标记物与阳性(CD49d)或阴性(CCR4)标志物与群体产生效应细胞因子的相对能力相关(图7D)。流式细胞术分析显示这些标志物表现出互斥的表达模式(图7E),eTreg细胞群中产生IL-2或IL-17的少数细胞在表达CD49d的细胞中最为突出,这和已有的报道结果一致。然而,仅CD49d-CCR4 +表型完全排除了完全产生效应细胞因子的eTreg细胞(图7G)。在CD127-Fr.III群体中观察到类似的趋势,其中CD49d-CCR4 +群体中不存在产生IL-17和IFN-g的细胞。然而,该群体在产生IL-2的能力方面仍然不同于相应的CD49d-CCR4 + eTreg细胞亚群(图7G)。FOXP3的相对蛋白水平与每个群体中产生细胞因子的细胞的比例成反比。因此,FOXP3表达逐渐从eTreg细胞(最高)降低至Fr.III CD127-和Fr.III CD127 +(最低,但仍然高于mTconv细胞)。此外,在每个群体中,CCR4+CD49d-细胞总是表达比CCR4-CD49d +细胞更高的FOXP3。最后,TSDR仅在eTreg细胞中完全去甲基化,并且主要在CD127-CCR4+CD49d-Fr.III细胞中甲基化,其仅产生IL-2,但不产生其他炎性细胞因子。因此,作者的蛋白质组学数据使其发现CCR4和CD49d区分具有不同能力的细胞,以在eTreg细胞和Fr.III群体内产生效应细胞因子。这些标记物与常用的Treg细胞标记物组合的组合可用于细胞纯化和诊断目的。例如,它们有助于阐明肿瘤中FOXP3+ CD4+T细胞的存在与患者愈后之间的关联,特别是对于癌症如结肠癌等。 文章小结
文章小结
通过对人类调节和常规CD4+T(Tconv)细胞的各种群体进行蛋白质组学和转录组学,获得调节性T(Treg)细胞特性的分子特征,鉴定并定义了所有Treg细胞的蛋白质表达特征,以及定义效应Treg细胞的独特特征。发现了Treg细胞中的代谢特征,以及细胞因子、TCR和共刺激受体信号传导途径的特异性适应性:适应——选择性STAT4缺陷——保护Treg细胞稳态模型,该途径通过炎性细胞因子起作用,而这些信号仍然可以通过其他途径诱发关键的转录因子和引导受体。此外,作者的研究揭示了识别具有不同功能特性的FOXP3+ CD4+T细胞的表面标志物。研究结果表明,信号通路中的适应性保护Treg细胞,并为进一步研究Treg细胞生物学提供了资源。
参考文献
1. Vogel, C., and Marcotte, E.M. (2012). Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 13, 227–232.
2. Arpaia, N., Green, J.A., et al . (2015). A distinct function of regulatory T cells in tissue protection. Cell 162, 1078–1089.
3. De Rosa, V., Galgani, M., et al. (2015). Glycolysis controls the induction of human regulatory T cells by modulating the expression of FOXP3 exon 2 splicing variants. Nat. Immunol16, 1174–1184.
4. Oh, H., Grinberg-Bleyer, Y., et al. (2017). An NF-kB transcription-factor-dependent lineage-specific transcriptional program promotes regulatory T cell identity and function. Immunity 47, 450–465.e5. -
利用iTRAQ研究光控细菌代谢用于肿瘤治疗
2018-06-25 16:56:45
题目:Optically-controlled bacterial metabolite for cancer therapy. Nature Communications IF 12.124 iTRAQ研究背景随着对共生微生物认识的日益加深,人们逐渐意识到肿瘤内部是细菌和细胞共生的复杂微环境,并且相当一部分的细菌可以特异性地从循环系统中富集到肿瘤部位。在先前的研究中,科研人员也发现这些共生细菌在肿瘤的发展过程中起到关键性作用。在人为改造下,这一类共生细菌能够富集到肿瘤部位,抑制肿瘤生长,从而为肿瘤治疗提供全新方案。近来,许多纳米级光催化材料如CdS和C3N4因其光电转换能力引起了广泛的关注,其可以将光能不断转换成电能。这些材料不仅能够增加细菌的代谢活性,而且能够避开负载极限,实现抗癌药物的富产。受上述光诱导还原的启发,本文作者将光催化系统与肿瘤靶向细菌融合,从而获得用于光控制的一氧化氮(NO)生成的生物/非生物混合物。首先合成了抑制自由基生成能力的碳量子点掺杂的碳化氮(CCN),以实现原位光电转换。然后,通过静电相互作用组装CCN和大肠杆菌(E. coli)以获得CCN@E. coli。本文中,作者提出了一个利用修饰的CCN@E. coli的光控细菌代谢物疗法(PMT)的概念。该技术利用修饰的CCN@E. coli将NO3-转化为在光照下具有癌症治疗效果的抗肿瘤一氧化氮(NO)。作者还利用iTRAQ定量蛋白质组学技术来研究其详细机制。PMT疗法优化了生物/非生物混合系统在生物医学中应用,转变了细菌癌症治疗方式。研究内容及结果1. PMT系统的特性和机理CCN@E. coli的制备如图1a所示。CCN通过静电相互作用与E. coli组装到一起。TEM图像显示修饰后,CCNs位于E. coli表面,在此过程中E. coli的形态没有发生变化(图1c)。如图1d所示,CCN和E. coli之间的空间重叠系数(0.94)也表明CCN@E. coli组装成功。作者推测,由于其合适的带隙,从CCN激发的光电子能够通过电子载体(例如NADH)转移至大肠杆菌的NO合成酶中。随后,NO3-可能以NADH依赖性方式酶促还原为NO(图1g)。因此作者利用经典的Griess方法研究了CCN@E. coli光学控制的NO释放行为(图2a)。在正常生理条件下,L-精氨酸通过一氧化氮合成酶阳性细胞如巨噬细胞转化为NO。然而,由于来源有限,并且NO可能最终被氧化成无毒的NO3-,所以生理条件下产生的NO的抗癌潜能力很微弱。在CCN@E.coli辅助下的PMT疗法可将不可逆的肿瘤内NO代谢转化为循环反应,最大限度地提高NO的生物利用度。PMT系统的机制是一个两步过程:首先,E.coli利用发光的CCN光电产生的电子进行内源性NO3 -还原和NO生成(图2g),然后,由产生的NO引发细胞凋亡。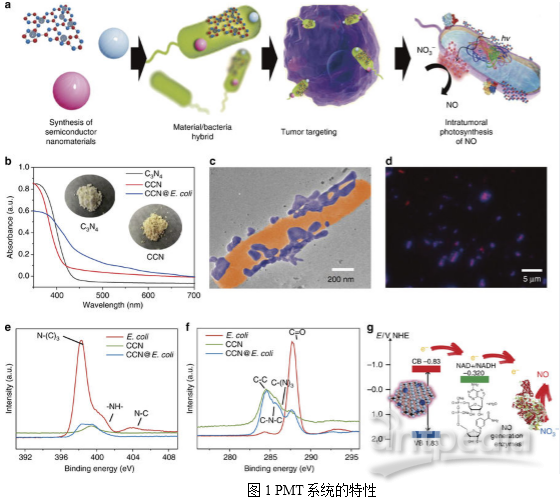 2. PMT的体外抗癌研究为了使癌细胞和细菌在不直接接触的情况下实现代谢物的交换,作者使用3D打印技术制作了一种共培养装置。在这种培养体系中,NO可以通过CCN@E. coli(装置外培养)的多孔膜扩散到癌细胞(装置内培养),而CCN@E. coli或癌细胞都不能通过这个膜迁移。然后利用二氨基荧光素-FM二乙酸酯(DAF-FM DA)测定细胞内NO的浓度。如图2i所示,4T1细胞中绿色荧光的增加表明,PMT系统产生的NO能有效扩散到附近的腔室,上调癌细胞中的NO的水平。没有光照射的野生型大肠杆菌和CCN@E. coli不能使细胞内NO浓度增加(图2j)。如图2k所示,使用CCN@E. coli光辐射的细胞存活率显著降低,在CCN@E. coli光辐射量为108 CFU/mL时,高达70%的4T1细胞能在24小时内被杀死。此外,作者还发现血红蛋白(一种NO清除剂)和Ac-DEVD-CHO(一种凋亡抑制剂)能有效地阻碍4T1细胞的凋亡(图2I)。基于上述观察,作者推断NO诱导的细胞凋亡是4T1细胞死亡的主要原因。
2. PMT的体外抗癌研究为了使癌细胞和细菌在不直接接触的情况下实现代谢物的交换,作者使用3D打印技术制作了一种共培养装置。在这种培养体系中,NO可以通过CCN@E. coli(装置外培养)的多孔膜扩散到癌细胞(装置内培养),而CCN@E. coli或癌细胞都不能通过这个膜迁移。然后利用二氨基荧光素-FM二乙酸酯(DAF-FM DA)测定细胞内NO的浓度。如图2i所示,4T1细胞中绿色荧光的增加表明,PMT系统产生的NO能有效扩散到附近的腔室,上调癌细胞中的NO的水平。没有光照射的野生型大肠杆菌和CCN@E. coli不能使细胞内NO浓度增加(图2j)。如图2k所示,使用CCN@E. coli光辐射的细胞存活率显著降低,在CCN@E. coli光辐射量为108 CFU/mL时,高达70%的4T1细胞能在24小时内被杀死。此外,作者还发现血红蛋白(一种NO清除剂)和Ac-DEVD-CHO(一种凋亡抑制剂)能有效地阻碍4T1细胞的凋亡(图2I)。基于上述观察,作者推断NO诱导的细胞凋亡是4T1细胞死亡的主要原因。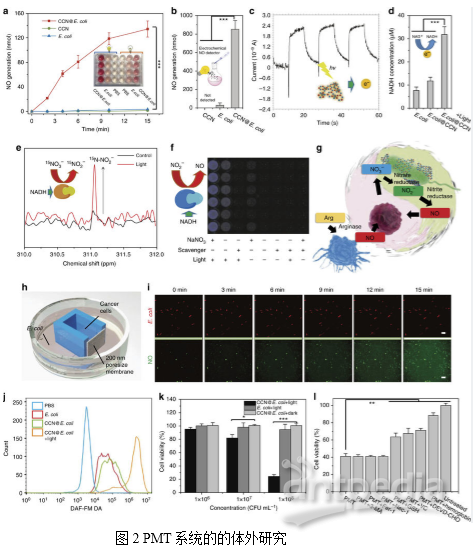 3. PMT系统的体内生物分布研究为评估PMT系统的肿瘤靶向能力,将DIR标记的CCN@E.coli静脉注射到4T1荷瘤小鼠。如图3a所示,随着时间的延长,在肿瘤位置内检测到荧光的也逐渐增强。如图3b所示,离体荧光成像显示大量的细菌积聚在肿瘤位置,而肝脏或肾脏滞留的细菌几乎可忽略不计。值得注意的是,CCN的修饰并不影响E.coli的肿瘤靶向能力(图3c)。通常,大多数合成药物载体由于其扩散限制,几乎不能穿透肿瘤组织。而作者推测PMT系统可能能够到达移植瘤的较深区域。结果表明,不同深度的肿瘤组织切片显示PMT系统分布于肿瘤组织中(图3d),并且发现移植瘤的中心聚积的细菌更多。为了全面揭示CCN@E.coli的肿瘤穿透性特征。作者使用表面活性剂辅助组织清除技术,即CLARITY,将肿瘤组织转化为透明的形式(图3e)。然后使用与肿瘤缺氧高度相关的生物标志物碳酸酐酶-Ⅸ(CA-9)对肿瘤缺氧区进行染色。从3D肿瘤荧光图像可观察到来自CCN@E.coli的红色荧光和来自CA-9的绿色荧光的共定位,而CCN@E.coli荧光可以在含氧量正常的区域发现(图3f)。进而证明了PMT系统可以充分利用大肠杆菌缺氧介导的趋化性,并且达到其他常规载体无法高效利用的缺氧区域。随后,作者研究了CCN@E.coli在主要代谢器官如肝、脾和肾中的清除动力学。通过PCR和定性免疫荧光染色证明大部分细菌可以随着的时间延长而被清除。3周后,在免疫活性小鼠中可以检测到极少量的细菌(图3h),血液生化和血液学分析未发现PMT的长期副作用(图3i)。
3. PMT系统的体内生物分布研究为评估PMT系统的肿瘤靶向能力,将DIR标记的CCN@E.coli静脉注射到4T1荷瘤小鼠。如图3a所示,随着时间的延长,在肿瘤位置内检测到荧光的也逐渐增强。如图3b所示,离体荧光成像显示大量的细菌积聚在肿瘤位置,而肝脏或肾脏滞留的细菌几乎可忽略不计。值得注意的是,CCN的修饰并不影响E.coli的肿瘤靶向能力(图3c)。通常,大多数合成药物载体由于其扩散限制,几乎不能穿透肿瘤组织。而作者推测PMT系统可能能够到达移植瘤的较深区域。结果表明,不同深度的肿瘤组织切片显示PMT系统分布于肿瘤组织中(图3d),并且发现移植瘤的中心聚积的细菌更多。为了全面揭示CCN@E.coli的肿瘤穿透性特征。作者使用表面活性剂辅助组织清除技术,即CLARITY,将肿瘤组织转化为透明的形式(图3e)。然后使用与肿瘤缺氧高度相关的生物标志物碳酸酐酶-Ⅸ(CA-9)对肿瘤缺氧区进行染色。从3D肿瘤荧光图像可观察到来自CCN@E.coli的红色荧光和来自CA-9的绿色荧光的共定位,而CCN@E.coli荧光可以在含氧量正常的区域发现(图3f)。进而证明了PMT系统可以充分利用大肠杆菌缺氧介导的趋化性,并且达到其他常规载体无法高效利用的缺氧区域。随后,作者研究了CCN@E.coli在主要代谢器官如肝、脾和肾中的清除动力学。通过PCR和定性免疫荧光染色证明大部分细菌可以随着的时间延长而被清除。3周后,在免疫活性小鼠中可以检测到极少量的细菌(图3h),血液生化和血液学分析未发现PMT的长期副作用(图3i)。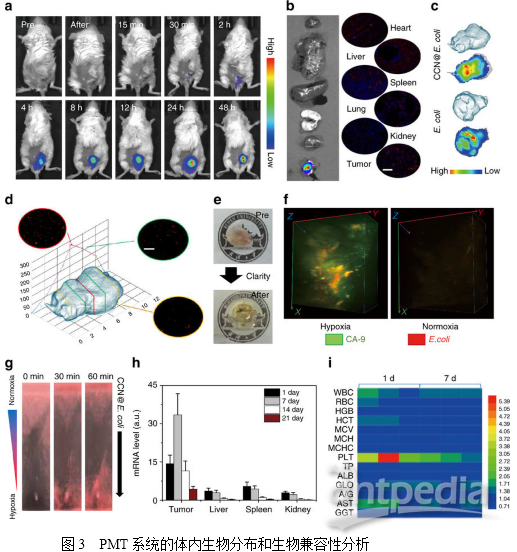 4. PMT系统中体内NO的生成为了进一步阐明细胞光合行为,作者设计并构建了Nrf2受控萤光素酶表达质粒用以探测体内NO的产生,之后转染4T1细胞以构建4T1Nrf2细胞用于原位NO的检测。如图4a所示,4T1Nrf2-luc细胞暴露在CCN@E.coli环境下会导致其生物发光快速增加,作者还研究了4T1Nrf2-luc肿瘤小鼠中PMT系统的体内效率。如图4b所示,单独的CCN和E.coli都不能诱导产生可检测的NO。而CCN@E.coli处理组的生物发光强度显著增加。接下来,作者还合成了用于NO检测的磁共振成像(MRI)探针Fe-MGD。如图4c所示,作者发现在注射了CCN@E.coli和光照后,小鼠肿瘤内的T1信号增强,进一步证明了体内光控制的NO生成。
4. PMT系统中体内NO的生成为了进一步阐明细胞光合行为,作者设计并构建了Nrf2受控萤光素酶表达质粒用以探测体内NO的产生,之后转染4T1细胞以构建4T1Nrf2细胞用于原位NO的检测。如图4a所示,4T1Nrf2-luc细胞暴露在CCN@E.coli环境下会导致其生物发光快速增加,作者还研究了4T1Nrf2-luc肿瘤小鼠中PMT系统的体内效率。如图4b所示,单独的CCN和E.coli都不能诱导产生可检测的NO。而CCN@E.coli处理组的生物发光强度显著增加。接下来,作者还合成了用于NO检测的磁共振成像(MRI)探针Fe-MGD。如图4c所示,作者发现在注射了CCN@E.coli和光照后,小鼠肿瘤内的T1信号增强,进一步证明了体内光控制的NO生成。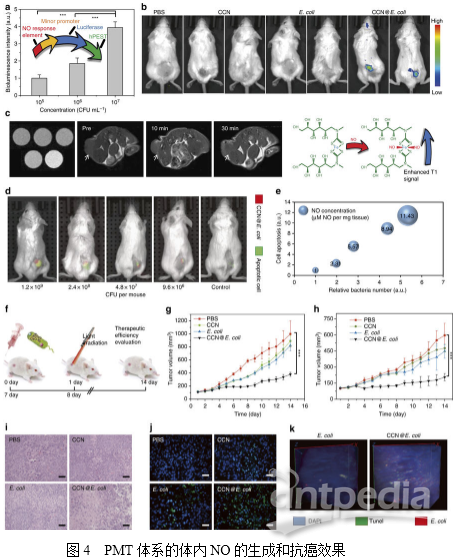 5. PMT的体内抗癌机制作者利用缀合有Cy5.5的膜联蛋白V29来研究PMT的体内治疗反应。如图4d所示,随着CCN@E.coli注射剂量的增加,CCN@E.coli在肿瘤部位的累积增多。图4e的半定量分析结果也证明了肿瘤内细菌数目、肿瘤细胞毒性和NO浓度呈正相关。此外,作者在4T1荷瘤小鼠和CT26荷瘤小鼠中检查了PMT的抗癌效率,治疗时间表如图4f所示。在4T1荷瘤小鼠中,PMT治疗抑制了79.3%的肿瘤生长(图4g)。而在CT26荷瘤小鼠中,PMT治疗抑制了70.2%的肿瘤生长(图4h)。这些结果表明PMT应该能够适用于治疗不同类型的癌症。随后,作者利用定量蛋白质组学技术探索了PMT的详细抗癌机制。利用iTRAQ技术从4T1荷瘤小鼠癌组织(PMT处理组和PBS对照组)中共鉴定到4735个蛋白。如图5a所示,在PMT治疗后,肿瘤组织内鉴定到222个上调的差异蛋白和17个下调的差异蛋白(倍数变化≥1.5和P <0.05,超几何检验)。聚类分析和主成分分析结果显示PBS处理组和PMT处理组之间存在显著差异。对差异蛋白进行GO分析发现,刺激应答、信号、细胞死亡、免疫系统和细胞杀伤相关的蛋白质在PMT组中显著上调(图5b)。相反,与细胞增殖和生长相关的蛋白更多地发生下调。这些数据表明PMT在肿瘤中引起高应激反应,这主要是由于NO诱导的氧化损伤,并且与抗氧化活性相关的蛋白质也显著降低。通常,NO对癌细胞的细胞毒性主要归因于其诱导氧化应激和引发DNA损伤的能力。图5c所示的维恩图表明,大肠杆菌感染/侵入相关蛋白与DNA损伤/氧化应激相关蛋白之间没有重叠。因此,蛋白质组学研究表明CCN@E.coli对癌细胞的直接细胞毒性主要归因于NO本身。有趣的是,在GO分析中发现了参与免疫应答的蛋白质水平显著升高。这一现象表明免疫系统可能有助于体内PMT的抗癌作用。PMT治疗后蛋白相互作用网络分析也可得出同样的结论。如图5e所示,KEGG分析发现抗原递呈途径被全面激活,并且证实了HMGB蛋白(一种参与触发抗原呈递的主导蛋白导致免疫原性细胞死亡)的上调。这些发现揭示了PMT可能通过HMGB引发MHC I类介导的途径诱导免疫原性细胞死亡。在体外模拟环境中,发现PMT治疗能够诱导DC突变(图5f)。这些结果表明,除了直接产生细胞毒性NO外,免疫应答也可能在在PMT的抗癌机制中起关键作用。
5. PMT的体内抗癌机制作者利用缀合有Cy5.5的膜联蛋白V29来研究PMT的体内治疗反应。如图4d所示,随着CCN@E.coli注射剂量的增加,CCN@E.coli在肿瘤部位的累积增多。图4e的半定量分析结果也证明了肿瘤内细菌数目、肿瘤细胞毒性和NO浓度呈正相关。此外,作者在4T1荷瘤小鼠和CT26荷瘤小鼠中检查了PMT的抗癌效率,治疗时间表如图4f所示。在4T1荷瘤小鼠中,PMT治疗抑制了79.3%的肿瘤生长(图4g)。而在CT26荷瘤小鼠中,PMT治疗抑制了70.2%的肿瘤生长(图4h)。这些结果表明PMT应该能够适用于治疗不同类型的癌症。随后,作者利用定量蛋白质组学技术探索了PMT的详细抗癌机制。利用iTRAQ技术从4T1荷瘤小鼠癌组织(PMT处理组和PBS对照组)中共鉴定到4735个蛋白。如图5a所示,在PMT治疗后,肿瘤组织内鉴定到222个上调的差异蛋白和17个下调的差异蛋白(倍数变化≥1.5和P <0.05,超几何检验)。聚类分析和主成分分析结果显示PBS处理组和PMT处理组之间存在显著差异。对差异蛋白进行GO分析发现,刺激应答、信号、细胞死亡、免疫系统和细胞杀伤相关的蛋白质在PMT组中显著上调(图5b)。相反,与细胞增殖和生长相关的蛋白更多地发生下调。这些数据表明PMT在肿瘤中引起高应激反应,这主要是由于NO诱导的氧化损伤,并且与抗氧化活性相关的蛋白质也显著降低。通常,NO对癌细胞的细胞毒性主要归因于其诱导氧化应激和引发DNA损伤的能力。图5c所示的维恩图表明,大肠杆菌感染/侵入相关蛋白与DNA损伤/氧化应激相关蛋白之间没有重叠。因此,蛋白质组学研究表明CCN@E.coli对癌细胞的直接细胞毒性主要归因于NO本身。有趣的是,在GO分析中发现了参与免疫应答的蛋白质水平显著升高。这一现象表明免疫系统可能有助于体内PMT的抗癌作用。PMT治疗后蛋白相互作用网络分析也可得出同样的结论。如图5e所示,KEGG分析发现抗原递呈途径被全面激活,并且证实了HMGB蛋白(一种参与触发抗原呈递的主导蛋白导致免疫原性细胞死亡)的上调。这些发现揭示了PMT可能通过HMGB引发MHC I类介导的途径诱导免疫原性细胞死亡。在体外模拟环境中,发现PMT治疗能够诱导DC突变(图5f)。这些结果表明,除了直接产生细胞毒性NO外,免疫应答也可能在在PMT的抗癌机制中起关键作用。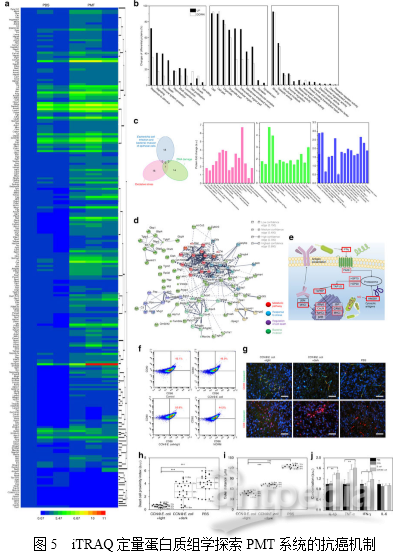 文章小结一氧化氮(NO)在浓度较高的环境可以引发肿瘤细胞凋亡。为了实现对细菌合成NO能力的提升及控制,作者将碳量子点掺杂的碳化氮(CCN)负载到大肠杆菌(E.coli)MG1655上,利用合成材料良好的光催化性能,从而提高NO的产生,这一方法可以很好地实现细菌在肿瘤富集。本文作者全面研究了光控细菌代谢疗法(PMT)中NO的生成,细胞毒性细胞杀伤作用及其相关机制。动物实验证明,该疗法在小鼠肿瘤模型上表现出了约80%的抑瘤率。这一发现对哺乳动物-微生物的共生关系有了更深认识,并且极大丰富现有肿瘤疗法的内涵。定量蛋白质组学结果也表明免疫反应可能与该疗法相关, PMT疗法可能对癌症免疫疗法具有促进作用。该研究策略将为活体生物材料的设计和制备提供一种新的方法和视角。参考文献1. Treweek, J. B. et al. Whole-body tissue stabilization and selective extractions via tissue-hydrogel hybrids for high-resolution intact circuit mapping and phenotyping. Nat. Protoc. 10, 1860–1896 (2015).2. Luo, C. H., Huang, C. T., Su, C. H. & Yeh, C. S. Bacteria-mediated hypoxiaspecific delivery of nanoparticles for tumors imaging and therapy. Nano Lett.16, 3493–3499 (2016).3. Zheng, J. H. et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med. 9, eaak9537 (2016).4. Zitvogel, L., Daillere, R., Roberti, M. P., Routy, B. & Kroemer, G. Anticancer effects of the microbiome and its products. Nat. Rev. Microbiol. 15, 465–478 (2017).5. Wang, B. et al. Enhanced biological hydrogen production from Escherichia coli with surface precipitated cadmium sulfide nanoparticles. Adv. Energy Mater. 7, 1700611 (2017).
文章小结一氧化氮(NO)在浓度较高的环境可以引发肿瘤细胞凋亡。为了实现对细菌合成NO能力的提升及控制,作者将碳量子点掺杂的碳化氮(CCN)负载到大肠杆菌(E.coli)MG1655上,利用合成材料良好的光催化性能,从而提高NO的产生,这一方法可以很好地实现细菌在肿瘤富集。本文作者全面研究了光控细菌代谢疗法(PMT)中NO的生成,细胞毒性细胞杀伤作用及其相关机制。动物实验证明,该疗法在小鼠肿瘤模型上表现出了约80%的抑瘤率。这一发现对哺乳动物-微生物的共生关系有了更深认识,并且极大丰富现有肿瘤疗法的内涵。定量蛋白质组学结果也表明免疫反应可能与该疗法相关, PMT疗法可能对癌症免疫疗法具有促进作用。该研究策略将为活体生物材料的设计和制备提供一种新的方法和视角。参考文献1. Treweek, J. B. et al. Whole-body tissue stabilization and selective extractions via tissue-hydrogel hybrids for high-resolution intact circuit mapping and phenotyping. Nat. Protoc. 10, 1860–1896 (2015).2. Luo, C. H., Huang, C. T., Su, C. H. & Yeh, C. S. Bacteria-mediated hypoxiaspecific delivery of nanoparticles for tumors imaging and therapy. Nano Lett.16, 3493–3499 (2016).3. Zheng, J. H. et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med. 9, eaak9537 (2016).4. Zitvogel, L., Daillere, R., Roberti, M. P., Routy, B. & Kroemer, G. Anticancer effects of the microbiome and its products. Nat. Rev. Microbiol. 15, 465–478 (2017).5. Wang, B. et al. Enhanced biological hydrogen production from Escherichia coli with surface precipitated cadmium sulfide nanoparticles. Adv. Energy Mater. 7, 1700611 (2017). -
iTRAQ蛋白质组揭示油菜卷叶机理
2018-06-25 16:18:55
文献:Histological, Physiological, and Comparative Proteomic Analyses Provide Insights into Leaf Rolling in Brassica napus.Journal of Proteome Research IF4.268 iTRAQ研究背景叶片是大多数植物光合作用的主要器官,叶片发育影响作物产量和植物结构。适度的卷叶被认为是作物理想型育种的一个重要组成部分,它能够改变植物的结构,提高光合效率,延缓叶片衰老,减轻干旱、高温和高光等胁迫带来的损害。近年来,在拟南芥和水稻等物种中,已经分离出了卷曲叶片突变体,并对它们的分子机制进行了深入的研究。油菜作为世界上最重要的油料作物之一,虽然也发现了一些卷曲叶片的突变体,但是对于其作用机制仍有待进一步的研究。本文作者主要利用iTRAQ定量蛋白质组学技术分析油菜卷叶突变株Bndcl1与野生株(WT)的差异蛋白,阐明了卷叶机理及其对植物的生理影响,有望促进油菜的理想株型育种。研究内容及结果1. 本研究中,作者选取了油菜卷叶突变株Bndcl1,首先经组织学对比发现,相比于野生株(WT),突变株(Bndcl1)幼苗期的叶片表型上比较紧凑(图 1A和B)。组织切片结果中发现Bndcl1叶绿体数目明显增多,特别是在海绵状的叶肉细胞中(图1C和D)。此外,作者发现,在Bndcl1突变体中,在远轴端韧皮部组织细胞较少,而在其叶脉以下的远轴端表皮细胞明显增大(图1E和F)。 2. 为了评估卷曲叶片对Bndcl1突变株光合性能的影响,作者测定了Chl的含量、Chl荧光和气体交换参数。与野生株相比,Bndcl1株中的叶绿体数量、PSII和净光合速率的有效量子产量显著增加。3. 活性氧(ROS)的生成在光合作用的光反应过程中是不可避免的,PSII极易受到光破坏的影响。光抑制的光合作用不可避免的损失会因ROS的产生而加剧,而ROS的产生会减慢PSII的修复。为了减少光氧化伤害,光合生物体已经进化出多种机制,而对抗氧化剂的上调是调节机制的一部分。因此,作者检测了Bndcl1和WT叶子的酶活性、脂质过氧化和ROS水平,发现在Bndcl1叶片中O2• − 和H2O2较低,由此说明其ROS水平也将明显降低(图2A和B)。此外,TBARS在突变株的低水平同样说明其膜脂过氧化的水平降低(图2C)。作者还检测比较了两者的SOD、POD和CAT酶活性,发现相比于WT株,在Bndcl1叶片中此酶活性显著增加。由此说明Bndcl1的光系统能更好的避免受到光氧化伤害。
2. 为了评估卷曲叶片对Bndcl1突变株光合性能的影响,作者测定了Chl的含量、Chl荧光和气体交换参数。与野生株相比,Bndcl1株中的叶绿体数量、PSII和净光合速率的有效量子产量显著增加。3. 活性氧(ROS)的生成在光合作用的光反应过程中是不可避免的,PSII极易受到光破坏的影响。光抑制的光合作用不可避免的损失会因ROS的产生而加剧,而ROS的产生会减慢PSII的修复。为了减少光氧化伤害,光合生物体已经进化出多种机制,而对抗氧化剂的上调是调节机制的一部分。因此,作者检测了Bndcl1和WT叶子的酶活性、脂质过氧化和ROS水平,发现在Bndcl1叶片中O2• − 和H2O2较低,由此说明其ROS水平也将明显降低(图2A和B)。此外,TBARS在突变株的低水平同样说明其膜脂过氧化的水平降低(图2C)。作者还检测比较了两者的SOD、POD和CAT酶活性,发现相比于WT株,在Bndcl1叶片中此酶活性显著增加。由此说明Bndcl1的光系统能更好的避免受到光氧化伤害。 4. 作者应用iTRAQ定量蛋白质组学技术研究Bndcl1与WT叶片蛋白质表达水平的差异,总共鉴定到5019个蛋白,差异蛋白有943个,其中发生上调的蛋白有451个,发生下调的蛋白有492个。作者对这些蛋白进行了GO、COG、KEGG等注释分析。通过对差异蛋白中发生上调和下调的蛋白分别进行GO分析并对比发现,在“细胞增殖”、“生物过程的调节”、“色素沉着”等功能方面,下调的蛋白数目要比上调的蛋白数多,具有抗氧化活性的功能的上调蛋白数目是下调蛋白数目的3倍多(图3)。COG分析表明,大多数下调蛋白参与翻译、核糖体结构和生物发生,而上调蛋白主要参与碳水化合物的运输和代谢、转录翻译后修饰、蛋白质的周转、分子伴侣和能量的产生和转化(图4)。KEGG 注释结果显示,尽管在“代谢途径”、“次生代谢产物的生物合成”和“淀粉和蔗糖代谢”中均有上调和下调的蛋白质参与,但在不同的途径中也涉及到不同差异蛋白质(图5)。
4. 作者应用iTRAQ定量蛋白质组学技术研究Bndcl1与WT叶片蛋白质表达水平的差异,总共鉴定到5019个蛋白,差异蛋白有943个,其中发生上调的蛋白有451个,发生下调的蛋白有492个。作者对这些蛋白进行了GO、COG、KEGG等注释分析。通过对差异蛋白中发生上调和下调的蛋白分别进行GO分析并对比发现,在“细胞增殖”、“生物过程的调节”、“色素沉着”等功能方面,下调的蛋白数目要比上调的蛋白数多,具有抗氧化活性的功能的上调蛋白数目是下调蛋白数目的3倍多(图3)。COG分析表明,大多数下调蛋白参与翻译、核糖体结构和生物发生,而上调蛋白主要参与碳水化合物的运输和代谢、转录翻译后修饰、蛋白质的周转、分子伴侣和能量的产生和转化(图4)。KEGG 注释结果显示,尽管在“代谢途径”、“次生代谢产物的生物合成”和“淀粉和蔗糖代谢”中均有上调和下调的蛋白质参与,但在不同的途径中也涉及到不同差异蛋白质(图5)。

 5. 为了进一步对这些差异蛋白的功能进行分类分析,作者进行了功能富集分析。通过对差异蛋白的富集分析结果发现,这些发生上调的蛋白能够改善Bndcl1株的光合性能和减轻氧化伤害,并且发现下调的蛋白参与叶片近-远轴极性建成。6. 作者最后从差异表达蛋白中挑选了多个蛋白进行了qRT-PCR表达验证。例如:与卷叶表型相关的基因PGY1, PGY2, PGY3, AE5, STV1 and SGS3,这几个基因的蛋白表达水平在蛋白质组学结果是显著下调。qRT-PCR验证结果显示,在Bndcl1株,这些基因mRNA表达水平显著降低,表明这些基因在蛋白质和mRNA水平上发生了一致的变化(图6A)。另外作者从转录水平上检测了叶极性的关键调节因子:AS1, AS2, KAN1, KAN2, REV, RDR6, AGO7, AGO1(图6B)。结果发现在Bndcl1株,AS1, KAN2发生了显著降低,AS2, KAN1没有显著变化,REV发生了显著下调。此外,RDR6, AGO7, AGO1也同样发生了显著下调。最后作者还对油菜甾醇相关的6个差异蛋白进行了转录水平的验证,结果与iTRAQ数据(图6C)一致,说明了定量蛋白质组学分析所获得的结果的可靠性。
5. 为了进一步对这些差异蛋白的功能进行分类分析,作者进行了功能富集分析。通过对差异蛋白的富集分析结果发现,这些发生上调的蛋白能够改善Bndcl1株的光合性能和减轻氧化伤害,并且发现下调的蛋白参与叶片近-远轴极性建成。6. 作者最后从差异表达蛋白中挑选了多个蛋白进行了qRT-PCR表达验证。例如:与卷叶表型相关的基因PGY1, PGY2, PGY3, AE5, STV1 and SGS3,这几个基因的蛋白表达水平在蛋白质组学结果是显著下调。qRT-PCR验证结果显示,在Bndcl1株,这些基因mRNA表达水平显著降低,表明这些基因在蛋白质和mRNA水平上发生了一致的变化(图6A)。另外作者从转录水平上检测了叶极性的关键调节因子:AS1, AS2, KAN1, KAN2, REV, RDR6, AGO7, AGO1(图6B)。结果发现在Bndcl1株,AS1, KAN2发生了显著降低,AS2, KAN1没有显著变化,REV发生了显著下调。此外,RDR6, AGO7, AGO1也同样发生了显著下调。最后作者还对油菜甾醇相关的6个差异蛋白进行了转录水平的验证,结果与iTRAQ数据(图6C)一致,说明了定量蛋白质组学分析所获得的结果的可靠性。 文章小结这项结果研究表明,叶片近-远轴极性和油菜甾醇代谢及信号通路的缺陷是导致Bndcl1突变株叶子向下卷曲表型的原因,叶片的适度卷曲能够提高光吸收、能量转移和CO2固定效率,进而提高了突变株的光合性能。此外,在突变株中,参与PSII修复周期的抗氧化剂和蛋白质水平的升高可能会减少非生物胁迫的光合损失。这项研究为油菜卷叶机制及卷叶对植物生理的影响提供了新的见解。因此,卷叶突变株因具有紧凑的结构,能提高光合效率和增强抗逆性,这也是作物育种理想株型。参考文献1. Yang, C.; Xu, L.; et al. iTRAQ-based proteomics of sunflower cultivars differing in resistance to parasitic weed Orobanche cumana. Proteomics 2017, 17, 1700009.2. Liang, J.; Liu, B.; et al. Genetic variation and divergence of genes involved in leaf adaxial-abaxial polarity establishment in Brassica rapa. Front. Plant Sci. 2016, 7, 94.3. Husbands, A. Y.; Benkovics, A. H.; et al. The ASYMMETRIC LEAVES complex employs multiple modes of regulation to affect adaxial-abaxial patterning and leaf complexity. Plant Cell 2015, 27, 3321-3335.4. Jia, H.; Shao, M.; et al. Proteome dynamics and physiological responses to short-term salt stress in brassica napus leaves. Plos One 2015, 10, e0144808.5. Yamaguchi, T.; Nukazuka, A.; Tsukaya, H. Leaf adaxial-abaxial polarity specification and laminaoutgrowth: Evolution and development. Plant Cell Physiol. 2012, 53, 1180-1194.
文章小结这项结果研究表明,叶片近-远轴极性和油菜甾醇代谢及信号通路的缺陷是导致Bndcl1突变株叶子向下卷曲表型的原因,叶片的适度卷曲能够提高光吸收、能量转移和CO2固定效率,进而提高了突变株的光合性能。此外,在突变株中,参与PSII修复周期的抗氧化剂和蛋白质水平的升高可能会减少非生物胁迫的光合损失。这项研究为油菜卷叶机制及卷叶对植物生理的影响提供了新的见解。因此,卷叶突变株因具有紧凑的结构,能提高光合效率和增强抗逆性,这也是作物育种理想株型。参考文献1. Yang, C.; Xu, L.; et al. iTRAQ-based proteomics of sunflower cultivars differing in resistance to parasitic weed Orobanche cumana. Proteomics 2017, 17, 1700009.2. Liang, J.; Liu, B.; et al. Genetic variation and divergence of genes involved in leaf adaxial-abaxial polarity establishment in Brassica rapa. Front. Plant Sci. 2016, 7, 94.3. Husbands, A. Y.; Benkovics, A. H.; et al. The ASYMMETRIC LEAVES complex employs multiple modes of regulation to affect adaxial-abaxial patterning and leaf complexity. Plant Cell 2015, 27, 3321-3335.4. Jia, H.; Shao, M.; et al. Proteome dynamics and physiological responses to short-term salt stress in brassica napus leaves. Plos One 2015, 10, e0144808.5. Yamaguchi, T.; Nukazuka, A.; Tsukaya, H. Leaf adaxial-abaxial polarity specification and laminaoutgrowth: Evolution and development. Plant Cell Physiol. 2012, 53, 1180-1194. -
SWATH分析玉米蛋白质丰度、转录水平和组织多样性揭示其发育调控规律
2018-04-27 17:25:38
文献:An integrated analysis of protein abundance, transcript. level and tissue diversity to reveal developmental regulation of maize.Journal of Proteome Research(IF4.268)关键技术:SWATHSWATH分析玉米蛋白质丰度、转录水平和组织多样性揭示发育调控规律研究背景采用SWATH-MS技术定量分析四种玉米组织中的蛋白质:未成熟雌穗(V7阶段),未成熟雄穗(V8阶段),授粉后20天(20_DAP)幼胚和14日龄幼苗根,并进行蛋白组&转录组关联分析,鉴定组织特异性高表达基因和蛋白质,了解组织结构和器官发生的调节机制。研究内容1)数据分析和蛋白质特性合并来自四个组织样本的蛋白组学数据,作者通过SWATH共获得646,658个图谱。用不同的置信度阈值和错误发现率(FDR)来评估SWATH-MS的检测能力。发现。一定程度上较低阈值可鉴定到更多独特肽和蛋白质(图1A,1B)。在置信度0.85和FDR 0.05下,检测的多肽和蛋白质数量与使用更严格的阈值标准—置信度0.90和FDR 0.02相比,检测的多肽和蛋白数量稍有增加(图1A,1B),但差异蛋白质数量变化不大(图1D)。因此,最终选择置信度0.85和FDR 0.05阈值,因为该阈值能检测更多的蛋白质来扩增蛋白质组数据,而不会显著增加差异蛋白数量。共有117,184个unique谱图匹配到10,606个unique肽段,最后共鉴定到4,551个蛋白质(图1E)。蛋白质丰度在重复实验中重复性很好,相关系数在0.84-0.90之间(图S1)。在雌穗,雄穗,幼胚和幼根中分别鉴定到3916、3707、3702和2871种蛋白质,在这四种组织中共鉴定到2269种蛋白质(图1E)。大多数蛋白质(70%)被不少于2个unique谱图覆盖,主要由10至25个氨基酸组成(图S2A,S2B)。大约64%的蛋白质显示出> 5%的蛋白序列覆盖率度,84%的蛋白质的分子质量> 20kDa。 2)蛋白质组&转录组关联分析基于Doreen Ware实验室的转录组数据,及qTeller中下载的RNA-seq数据,作者对蛋白质组和转录组数据进行了关联分析。先将每个组织中的蛋白质丰度与相应的mRNA水平进行比较。然后过滤掉那些低mRNA水平(RPKM<1)的蛋白质,总共保留了4,314种(94.8%)蛋白,其中雌穗3554个,雄穗3404个,幼胚3417个和幼根2370个(图2A)。四个组织中共鉴定到2045个蛋白,表明大量检测到的蛋白存在于各个组织中。此外,还鉴定到了组织特异性高度表达的蛋白,其中幼胚253个,幼根181个,雌穗123个和雄穗43个(图2A)。其次,尽管蛋白质丰度与组织中的mRNA水平呈正相关,但Pearson相关系数很低,从0.35(幼根)到0.43(幼胚)(图2B),这表明转录水平并不总是与蛋白质丰度一一对应。去掉组织特异性高度表达的蛋白质,以变化倍数> 2倍和P值<0.01,FDR <0.05为筛选条件,共有3714个差异蛋白至少在两个组织中共同鉴定到。结果表明,在雌穗和雄穗之间鉴定到227个差异蛋白,在雌穗和幼根之间鉴定到799个差异蛋白(图2D-E)。差异蛋白质的数量反映了各组织形态和功能的差异。为了探究蛋白质与转录本丰度相关性较低的原因,作者将这些差异蛋白质分成如下三个亚组:i)正相关亚组(PC):mRNA水平与蛋白质丰度正相关;ii)低相关性亚组(LC):编码差异蛋白的转录本与蛋白质丰度相关但不显著;iii)负相关亚组(NC):mRNA水平与蛋白质丰度呈负相关。作者发现大约50%的差异蛋白质可以归类到PC亚组中(图2E),这表明过半的差异蛋白质的差异丰度主要由这些基因的表达差异所决定。然而,大约42%的差异蛋白,从31%到62%不等,归类到到LC亚组(图2E)。这种现象通常是由于RNA转录,加工和转换以及蛋白质翻译和转换的所导致的。4.9%至15%蛋白质归类到NC亚组(图2E),这表明转录组不能完全解释在组织或器官的不同功能和形态的分子基础上的蛋白质组,由此突出了蛋白质组学研究的重要性。
2)蛋白质组&转录组关联分析基于Doreen Ware实验室的转录组数据,及qTeller中下载的RNA-seq数据,作者对蛋白质组和转录组数据进行了关联分析。先将每个组织中的蛋白质丰度与相应的mRNA水平进行比较。然后过滤掉那些低mRNA水平(RPKM<1)的蛋白质,总共保留了4,314种(94.8%)蛋白,其中雌穗3554个,雄穗3404个,幼胚3417个和幼根2370个(图2A)。四个组织中共鉴定到2045个蛋白,表明大量检测到的蛋白存在于各个组织中。此外,还鉴定到了组织特异性高度表达的蛋白,其中幼胚253个,幼根181个,雌穗123个和雄穗43个(图2A)。其次,尽管蛋白质丰度与组织中的mRNA水平呈正相关,但Pearson相关系数很低,从0.35(幼根)到0.43(幼胚)(图2B),这表明转录水平并不总是与蛋白质丰度一一对应。去掉组织特异性高度表达的蛋白质,以变化倍数> 2倍和P值<0.01,FDR <0.05为筛选条件,共有3714个差异蛋白至少在两个组织中共同鉴定到。结果表明,在雌穗和雄穗之间鉴定到227个差异蛋白,在雌穗和幼根之间鉴定到799个差异蛋白(图2D-E)。差异蛋白质的数量反映了各组织形态和功能的差异。为了探究蛋白质与转录本丰度相关性较低的原因,作者将这些差异蛋白质分成如下三个亚组:i)正相关亚组(PC):mRNA水平与蛋白质丰度正相关;ii)低相关性亚组(LC):编码差异蛋白的转录本与蛋白质丰度相关但不显著;iii)负相关亚组(NC):mRNA水平与蛋白质丰度呈负相关。作者发现大约50%的差异蛋白质可以归类到PC亚组中(图2E),这表明过半的差异蛋白质的差异丰度主要由这些基因的表达差异所决定。然而,大约42%的差异蛋白,从31%到62%不等,归类到到LC亚组(图2E)。这种现象通常是由于RNA转录,加工和转换以及蛋白质翻译和转换的所导致的。4.9%至15%蛋白质归类到NC亚组(图2E),这表明转录组不能完全解释在组织或器官的不同功能和形态的分子基础上的蛋白质组,由此突出了蛋白质组学研究的重要性。 3)组织特异性高表达蛋白的功能分类基于蛋白质丰度作者在四种组织中共鉴定了600种组织特异性高表达蛋白质(图3A)。在细胞的整个蛋白质组中,蛋白质的低覆盖率是众所周知的,在所有现有的蛋白质组技术,包括SWATH-MS都有明显的缺陷。这种低覆盖率可能导致对组织特异性高表达蛋白的数量的过高估计。因此,作者检索了来自于qTeller的全长转录组数据和RNA-seq数据中编码600个组织特异性高表达蛋白的基因的表达水平,发现大部分基因(84.3%)在多种组织中都有表达,只有94种蛋白编码基因(15.7%)显示出组织特异性表达模式(图3A)。另外,26种蛋白质在雌穗和雄穗中共表达,称为花序特异性蛋白质。作者利用qRT-PCR交叉验证了18个基因的表达水平,15个为组织特异性表达基因,3个呈现出花序特异性表达模式(图3B,3D)。此外,作者还利用Western blot验证了2个幼胚特异性蛋白(GRMZM2G354013和GRMZM2G054916)(图3C,3E)。作者鉴定了94个组织特异性蛋白,其中幼根60个,幼胚27个,雌穗5个和雄穗2个。幼根特异性蛋白质直接或间接地与氧化还原过程和脱氢有关,包括12个过氧化物酶超家族蛋白,3个还原酶和8个水解酶活性蛋白,表明对氧化胁迫的响应在幼根的发育和生长中起重要作用。在27个幼胚特异性蛋白中,有5种植物脂质转移蛋白/种子储存相关蛋白质,3种氧化还原酶活性蛋白质和2种已知的与光形态相关的蛋白质转录因子(AtPIF1和AtSPT转录因子)。7个花序特异性蛋白与花序结构和花器官的发育高度相关。值得注意的是,有18种蛋白质在雌穗和雄穗中都有很高的丰度,但其mRNA水平在四个组织中都呈现出的低水平或没有被检测到。
3)组织特异性高表达蛋白的功能分类基于蛋白质丰度作者在四种组织中共鉴定了600种组织特异性高表达蛋白质(图3A)。在细胞的整个蛋白质组中,蛋白质的低覆盖率是众所周知的,在所有现有的蛋白质组技术,包括SWATH-MS都有明显的缺陷。这种低覆盖率可能导致对组织特异性高表达蛋白的数量的过高估计。因此,作者检索了来自于qTeller的全长转录组数据和RNA-seq数据中编码600个组织特异性高表达蛋白的基因的表达水平,发现大部分基因(84.3%)在多种组织中都有表达,只有94种蛋白编码基因(15.7%)显示出组织特异性表达模式(图3A)。另外,26种蛋白质在雌穗和雄穗中共表达,称为花序特异性蛋白质。作者利用qRT-PCR交叉验证了18个基因的表达水平,15个为组织特异性表达基因,3个呈现出花序特异性表达模式(图3B,3D)。此外,作者还利用Western blot验证了2个幼胚特异性蛋白(GRMZM2G354013和GRMZM2G054916)(图3C,3E)。作者鉴定了94个组织特异性蛋白,其中幼根60个,幼胚27个,雌穗5个和雄穗2个。幼根特异性蛋白质直接或间接地与氧化还原过程和脱氢有关,包括12个过氧化物酶超家族蛋白,3个还原酶和8个水解酶活性蛋白,表明对氧化胁迫的响应在幼根的发育和生长中起重要作用。在27个幼胚特异性蛋白中,有5种植物脂质转移蛋白/种子储存相关蛋白质,3种氧化还原酶活性蛋白质和2种已知的与光形态相关的蛋白质转录因子(AtPIF1和AtSPT转录因子)。7个花序特异性蛋白与花序结构和花器官的发育高度相关。值得注意的是,有18种蛋白质在雌穗和雄穗中都有很高的丰度,但其mRNA水平在四个组织中都呈现出的低水平或没有被检测到。 4)差异表达蛋白的功能分类GO富集确定差异蛋白功能类别。较其他组织对,雌穗和雄穗间鉴定到的差异蛋白更少,这些差异蛋白主要富集到“细胞分裂素介导的信号调节”,“乙酰辅酶A代谢过程”和“分生组织生长”(图4A),表明细胞分裂素介导的信号在雌穗和雄穗间的细胞分裂活性存在差异。雄穗中UB3蛋白质丰度比雌穗中高1.7倍,表明UB蛋白质积累的增加可能支持雄穗长枝的长出。雌穗和幼胚间的差异蛋白主要富集到“脂质生物合成过程”,“分生组织起始”和“生殖结构发育”条目。雄穗和幼胚间的差异蛋白也富集到这些GO条目(图4B,4C)。差异蛋白中不同GO类别反映了花序和幼胚间生理和发育上的差异。此外,雌穗/雄穗和幼根间的差异蛋白显著富集到“对无机物质的响应”和“有机酸生物合成过程”(图4D,4F)。幼胚和幼根间的差异蛋白主要富集到“有机酸代谢过程”,“对无机物质的反应”和“氧化还原”(p = 2.6 E-12)等。与“脂质生物合成过程”和“脂肪酸生物合成过程”有关的蛋白质在幼胚中显著富集,而涉及“响应无机物质”,“氧化还原”和“醇代谢过程”的蛋白质在幼根中显著富集。
4)差异表达蛋白的功能分类GO富集确定差异蛋白功能类别。较其他组织对,雌穗和雄穗间鉴定到的差异蛋白更少,这些差异蛋白主要富集到“细胞分裂素介导的信号调节”,“乙酰辅酶A代谢过程”和“分生组织生长”(图4A),表明细胞分裂素介导的信号在雌穗和雄穗间的细胞分裂活性存在差异。雄穗中UB3蛋白质丰度比雌穗中高1.7倍,表明UB蛋白质积累的增加可能支持雄穗长枝的长出。雌穗和幼胚间的差异蛋白主要富集到“脂质生物合成过程”,“分生组织起始”和“生殖结构发育”条目。雄穗和幼胚间的差异蛋白也富集到这些GO条目(图4B,4C)。差异蛋白中不同GO类别反映了花序和幼胚间生理和发育上的差异。此外,雌穗/雄穗和幼根间的差异蛋白显著富集到“对无机物质的响应”和“有机酸生物合成过程”(图4D,4F)。幼胚和幼根间的差异蛋白主要富集到“有机酸代谢过程”,“对无机物质的反应”和“氧化还原”(p = 2.6 E-12)等。与“脂质生物合成过程”和“脂肪酸生物合成过程”有关的蛋白质在幼胚中显著富集,而涉及“响应无机物质”,“氧化还原”和“醇代谢过程”的蛋白质在幼根中显著富集。 文章小结作者利用SWATH-MS技术在玉米4种组织中共鉴定到4551种蛋白质,与之前iTRAQ鉴定到的蛋白数目相当。基于蛋白质组和转录组关联分析,作者推断出蛋白质丰度与mRNA水平呈正相关,具有弱到中等的相关系数。然而,一些特定组织中与功能或结构相关的一些关键蛋白质在转录水平上被时空调控,在4种组织中不同蛋白表现出不同的表达模式。此外,组织特异性高表达的蛋白和差异蛋白能富集到不同GO类别中。组织特异性高度表达的蛋白质亚组通过交叉实验验证可作为潜在生物标志物。这加深了对玉米组织和器官发育分子机制的理解,并为其生物标志物的发现提供了全面的线索。备注:本文出于传递更多信息为目的解读该文献内容,不希望转载个人可与我联系,会立即删除处理。
文章小结作者利用SWATH-MS技术在玉米4种组织中共鉴定到4551种蛋白质,与之前iTRAQ鉴定到的蛋白数目相当。基于蛋白质组和转录组关联分析,作者推断出蛋白质丰度与mRNA水平呈正相关,具有弱到中等的相关系数。然而,一些特定组织中与功能或结构相关的一些关键蛋白质在转录水平上被时空调控,在4种组织中不同蛋白表现出不同的表达模式。此外,组织特异性高表达的蛋白和差异蛋白能富集到不同GO类别中。组织特异性高度表达的蛋白质亚组通过交叉实验验证可作为潜在生物标志物。这加深了对玉米组织和器官发育分子机制的理解,并为其生物标志物的发现提供了全面的线索。备注:本文出于传递更多信息为目的解读该文献内容,不希望转载个人可与我联系,会立即删除处理。 -
iTRAQ蛋白组学技术应用
2017-12-01 13:42:48
iTRAQ(isobaric tags for relative and absolute quantitation)技术是由AB SCIEX公司研发的一种体外同重同位素标记的相对与绝对定量技术,利用多种同位素试剂标记蛋白多肽N末端或赖氨酸侧链基团,经高精度质谱仪串联分析,可同时比较多达8种样品之间的蛋白表达量,是近年来定量蛋白质组学中应用最广泛的高通量筛选技术。那么iTRAQ试剂主要构成呢?iTRAQ试剂分为报告基团、平衡基团、氨基酸反应基团,报告基团共有8种(113、114、115、116、117、118、119、121Da),与平衡基团质量相加正好是305Da。氨基酸反应基团与肽链N端赖氨酸侧链共价链接,从而将报告基团和平衡基团标记到肽段上。在一级质谱中,不同来源的相同肽段被连接上总质量相同的iTRAQ标签试剂,具有相同的质荷比,表现为一个峰。在二级质谱中,iTRAQ标签试剂在不同基团连接处发生断裂,表现出不同质荷比的峰(平衡基团发生中性丢失)。根据波峰高度和面积,可以得到同一蛋白在不同样本中或在不同处理条件下的表达差异。 iTRAQ技术作为经典蛋白质组学定量技术,它的优势?灵敏度高:可检测出较低丰度蛋白,胞浆蛋白、膜蛋白、核蛋白、胞外蛋白等;分离能力强:可分离出酸/碱性蛋白,小于 10K 或大于200K的蛋白、难溶性蛋白;高通量:可同时对8个样本进行分析,特别适用于采用多种处理方式或来自多个处理时间的样本的差异蛋白分析;结果可靠准确:定性与定量同步进行,同时得出鉴定和定量结果,重复样品间的蛋白表达量相关性可达到 0.95 以上;自动化程度高:液质连用,自动化操作,分析速度快,分离效果好。除了涉及iTRAQ蛋白质组学实验样品、实验流程、质谱下机原始数据等,质谱数据分析也显得尤为重要,试想拿到数据却无法分析,无法得知实验最终结果是否与预期相符,可谓是万里长征只差一步。大数据时代的到来引起了高校对大数据分析关注,很多都陆续添加了生物信息学、生物统计学等课程,如果此刻的您也很感兴趣,不妨多深究下。iTRAQ蛋白质组学--生物信息学分析内容:iTRAQ鉴定:包括鉴定结果统计、重复性分析、Unique肽段数分布、肽段长度分布、蛋白覆盖度分布;iTRAQ定量:包括定量信息统计、蛋白质丰度比(FC)分布、表达量层次聚类分析;蛋白质功能注释:包括GO注释、COG注释和Pathway代谢通路注释;差异蛋白的富集分析:包括差异蛋白的GO富集分析、差异蛋白的Pathway富集分析。如果要做iTRAQ蛋白质组服务,需要注意什么呢?样本,外包公司销售会和您沟通的,不同样本由于高丰度/低丰度蛋白含量不同也会有差异的,还不如先让对方技术评估下实验方案。可能要强调一点,最后质谱下机原始数据需要结合数据库检索,所以要求物种具有蛋白质参考数据库、EST 序列(转录组)或基因组注释信息等,不清楚的,可以请外包公司帮您查下。iTRAQ现今发表文献网上有很多,可以下载学习,通过文献可以更清楚了解iTRAQ技术应用,也可@3268908524索取相关iTRAQ发表文献,加深对iTRAQ蛋白质组学的理解,拓宽项目实验设计思路。下期接着回归到非编码RNA上,也是小编对非编码RNA很感兴趣,就是这么任性!换个方向,也换个视野,知识都需要消化吸收,才能更好地加以利用。一本正经地说,噗嗤……
iTRAQ技术作为经典蛋白质组学定量技术,它的优势?灵敏度高:可检测出较低丰度蛋白,胞浆蛋白、膜蛋白、核蛋白、胞外蛋白等;分离能力强:可分离出酸/碱性蛋白,小于 10K 或大于200K的蛋白、难溶性蛋白;高通量:可同时对8个样本进行分析,特别适用于采用多种处理方式或来自多个处理时间的样本的差异蛋白分析;结果可靠准确:定性与定量同步进行,同时得出鉴定和定量结果,重复样品间的蛋白表达量相关性可达到 0.95 以上;自动化程度高:液质连用,自动化操作,分析速度快,分离效果好。除了涉及iTRAQ蛋白质组学实验样品、实验流程、质谱下机原始数据等,质谱数据分析也显得尤为重要,试想拿到数据却无法分析,无法得知实验最终结果是否与预期相符,可谓是万里长征只差一步。大数据时代的到来引起了高校对大数据分析关注,很多都陆续添加了生物信息学、生物统计学等课程,如果此刻的您也很感兴趣,不妨多深究下。iTRAQ蛋白质组学--生物信息学分析内容:iTRAQ鉴定:包括鉴定结果统计、重复性分析、Unique肽段数分布、肽段长度分布、蛋白覆盖度分布;iTRAQ定量:包括定量信息统计、蛋白质丰度比(FC)分布、表达量层次聚类分析;蛋白质功能注释:包括GO注释、COG注释和Pathway代谢通路注释;差异蛋白的富集分析:包括差异蛋白的GO富集分析、差异蛋白的Pathway富集分析。如果要做iTRAQ蛋白质组服务,需要注意什么呢?样本,外包公司销售会和您沟通的,不同样本由于高丰度/低丰度蛋白含量不同也会有差异的,还不如先让对方技术评估下实验方案。可能要强调一点,最后质谱下机原始数据需要结合数据库检索,所以要求物种具有蛋白质参考数据库、EST 序列(转录组)或基因组注释信息等,不清楚的,可以请外包公司帮您查下。iTRAQ现今发表文献网上有很多,可以下载学习,通过文献可以更清楚了解iTRAQ技术应用,也可@3268908524索取相关iTRAQ发表文献,加深对iTRAQ蛋白质组学的理解,拓宽项目实验设计思路。下期接着回归到非编码RNA上,也是小编对非编码RNA很感兴趣,就是这么任性!换个方向,也换个视野,知识都需要消化吸收,才能更好地加以利用。一本正经地说,噗嗤……


标题搜索
我的存档
数据统计
- 访问量: 0
- 日志数: 20
- 建立时间: 2017-11-09
- 更新时间: 2019-06-27
