上海欧易生物医学科技有限公司

 分享至
分享至



Cell新进展!人乳腺癌肿瘤及免疫生态系统的单细胞图谱


前言
乳腺癌是全球女性癌症死亡的主要原因,对乳腺癌生态系统的了解是实施精准医学的基础。单细胞基因组和转录组为乳腺癌的分析提供了肿瘤内基因组多样性和肿瘤间克隆组成差异的见解,但分析的细胞和肿瘤很少。鉴于乳腺癌细胞表型和细胞关系的异质性,患者分类和治疗应考虑整个肿瘤生态系统。
单细胞转录组测序研究为乳腺癌免疫细胞表型多样性和生态系统的进一步探索提供了可能,为探究大型患者队列的研究奠定了基础。因此研究者应用单细胞测序流式细胞仪对来自 144 个人类乳腺肿瘤样本的数百万个细胞进行分析,阐明了乳腺癌生态系统中的表型多样性和肿瘤免疫细胞关系。该单细胞图谱为基于乳腺癌生态系统的患者分类奠定了基础。
由苏黎世大学Bernd Bodenmiller教授课题组在Cell 期刊(IF:66.850)发表了题为 “A Single-Cell Atlas of the Tumor and Immune Ecosystem of Human Breast Cancer ”的研究成果,通过单细胞转录组、单细胞蛋白组,质谱流式等研究技术,描绘了免疫生态系统的单细胞图谱。为肿瘤治疗及其免疫环境的精准医学发展提供了理论依据。

研究思路

研究内容
1. 研究材料
144 个肿瘤样本、46 个近肿瘤样本和来自四个缩小乳房成形术的组织的 2600 万个细胞
2.研究方法
组织准备:手术切除后,新鲜组织样本立即转移到预冷的 MACS 组织储存溶液
抗体和抗体标记
抗体染色和细胞体积定量
免疫荧光成像
量化和统计分析
研究结果
1.乳腺癌生态系统的单细胞蛋白质组图谱
研究者对 144 个肿瘤样本进行了大规模单细胞蛋白质组分析,包括54 个luminal A,71个luminal B,6个 luminal B-HER2+,1个HER2+,和6个TN 肿瘤。组织病理学将样本分为 106 个浸润性导管、15 个浸润性小叶和 19 个混合或其他肿瘤。使用自动化系统从所有组织样本中生成单细胞悬浮液,添加标签后用于抗体染色,并同时通过质谱流式细胞仪进行分析(图 1 A 和S1 A)。分析产生了2600 万个单细胞图谱,每个样本平均有 84.7% 的活细胞、非凋亡细胞(图 S1B 和 S1C)。
为了确保高质量的数据,研究者确认了跨条形码板的重复样本的标记表达以及跨抗体组的活细胞和免疫细胞频率的相似性(图 S1 D 和 S1E)。


图S1 | 乳腺癌单细胞图谱的细胞类型鉴定
为了可视化肿瘤和非肿瘤细胞的多样性,作者使用降维算法 t 分布随机邻域嵌入生成了二维图。大多数细胞是上皮细胞或免疫细胞,内皮细胞和成纤维细胞的丰度较低(图 1B)。额外的成纤维细胞亚群和脂肪细胞可能在被描述为“其他”的细胞中(图1C)。为了比较肿瘤和非肿瘤组织之间的细胞类型频率,作者应用 PhenoGraph 算法,将数据划分为 42 个簇(图 S1 M 和 S1N)。与非肿瘤组织相比,乳腺肿瘤富含上皮细胞,并且含有更少的内皮细胞和成纤维细胞(图1D)。FAP+SMA+成纤维细胞在肿瘤中比在近肿瘤组织中更丰富(图 S1 O 和 S1P)。细胞类型频率在肿瘤亚型之间和之间有所不同,在 TN 和 HER2 +样本中观察到的免疫细胞频率高于其他乳腺癌类型(图 1 E 和S1 P)。
2.乳腺癌的免疫状况
T细胞和髓细胞是我们研究中最丰富的免疫细胞类型;较少检测到自然杀伤细胞(NK)、B细胞、粒细胞、浆细胞、嗜碱性细胞和浆细胞样树突状细胞(图2A和S2A S2D)。乳腺肿瘤富含T细胞和B细胞,与肿瘤旁组织相比,含有较低频率的NK细胞和粒细胞(图2A)。如前所述,肿瘤相关免疫细胞频率在患者间存在显著差异(图2A) 。



图2 | 乳腺癌的免疫状况
T 细胞和巨噬细胞可以发挥促肿瘤或抗肿瘤活性,通过 t-SNE 和 PhenoGraph 对 T 细胞的深入分析确定了十个 CD4 +和十个 CD8 + T 细胞簇(T01-T20;图 2 B-2D)。大多数 T 细胞簇具有效应记忆表型(CD197low, CD45RAlow),肿瘤相关 T 细胞作为表型连续体存在于 CD4 +和 CD8 +谱系中(图 2 D 和S2 E)。CD4 + T 细胞中 PD-1 的平均表达水平高于CD8 + T 细胞(图 2 F)。PD-1 的平均表达水平和 PD-1 + T 细胞频率在 CD4 +和 CD8 +隔室中相关,支持这些细胞由 T 细胞扩增产生的假设。
此外,研究者观察到 ER -和 ER +肿瘤的 T 细胞状况的差异,包括 ER -疾病中更高频率的 T-regs (图2H)。在超过一半的 ER -肿瘤但只有 12% 的 ER +肿瘤中,超过 10% 的 T 细胞表达 PD-1(图 S2 G)。不同的 PD-1 +表型分别富集:PD-1highCTLA-4 + CD38 +T 细胞在 ER -肿瘤中更常见,而 PD-1int CTLA-4 - CD38 - T 细胞(T14)在 ER +肿瘤中富集(图2H)。然而,许多 ER +肿瘤确实显示 PD-1高CTLA-4 + CD38 + T 细胞和 T-reg 的频率与 ER -肿瘤相当或更高(图2H)。


图S2 | 乳腺肿瘤和非肿瘤组织中的免疫细胞表型分析
ER +肿瘤可根据低增殖和高增殖分别分为luminal A和luminal B。超过 10% 的 T 细胞在 18% 的 luminal B 肿瘤中表达 PD-1,但只有 7% 的 luminal A 肿瘤(图 S2 H)。PD-1 int CTLA-4 - CD38 - T 细胞 (T07) 在 luminal A 疾病中更常见,而 T-regs 在 luminal B 肿瘤中富集(图 2 I)。作者还在不同级别的肿瘤中观察到不同的 T 细胞景观。在 28% 的 3 级肿瘤、9% 的 2 级肿瘤和 10% 的 1 级肿瘤中, PD-1 + T 细胞占 T 细胞的 10% 以上(图 S2 I)。3级肿瘤具有更多的PD-1高CTLA-4+ CD38 + T 细胞(T09 和 T11)和更少的 PD-1 int CTLA-4 - CD38 - T 细胞(T07 和 T14)比低级别肿瘤(图 S2 J)。这表明免疫抑制的 T 细胞景观与预后不良的肿瘤有关,包括 ER -、高增殖和高级别肿瘤,但也在一部分 ER +肿瘤中观察到。
3.乳腺肿瘤富含免疫抑制性巨噬细胞表型
为了表征TAM群体,将 t-SNE 和 PhenoGraph 应用于所有骨髓细胞(图 2 J 和S2 D),产生五类 19 个骨髓簇(M01-M19)。与近肿瘤组织相比,肿瘤富含 TAM,缺乏组织驻留巨噬细胞(M08 和 M09)、经典循环(M06)和促炎(M15)单核细胞(图2M)。TAM 的浸润与侵袭性疾病有关。
与 ER +肿瘤相比, ER -肿瘤包含更高频率的 M01 和 M17 PD-L1 + TAM,具有 M04、M05、M10 或 M12 表型的骨髓细胞更少(图 2 N)。一部分 ER +肿瘤具有 M01 和 M02 PD-L1 + TAM,其频率与 ER -肿瘤相当或更高(图 2 N 和S2 L)。与 Luminal A 肿瘤相比,Luminal B 肿瘤含有更多具有 M07 或 M17 表型的髓细胞,较少具有 M04 表型,以及更多的 PD-L1 + TAM。图 2 O 和S2 M)。与 2 级肿瘤相比,PD-L1 + TAM 在 3 级肿瘤中富集(图 S2 N)。与低级别肿瘤相比,3 级肿瘤包含更少的具有 M04 或 M05 表型的细胞,但更多的经典单核细胞 (M06)(图 S2 O)。
4.肿瘤上皮细胞异质性和表型异常
对来自肿瘤和非肿瘤组织的上皮细胞的分析(STAR 方法)揭示了上皮标志物的双峰和梯度样表达,表明许多不同的细胞表型(图3A和3B )。在 PhenoGraph 中实施的共识聚类方法(图 S3 A;STAR 方法)揭示了 45 个上皮细胞簇(Ep01-Ep45)。层次聚类根据标记表达将它们分为七个管腔组 L1-L7 和两个基础组 B1 和 B2(图 3 C、S3 B 和 S3C)。

图3 | 乳腺癌肿瘤细胞表型研究
研究者根据谱系标记表达模式鉴定了乳房成形术和近肿瘤组织中的管腔和肌上皮细胞(图 3 C、3D 和S3 D)。乳腺上皮细胞系证实了这些模式的可靠性(图 3 E)。鉴定了增殖的 (Ki-67 + ) 非肿瘤管腔细胞 (图 S3 E)。大约 55% 的肿瘤衍生细胞是 L1 和 L2 组的成员,这表明肿瘤样本中富含分化的正常样管腔细胞。L3-L7 组以肿瘤细胞为主(图 3C)。L3 组表型显示高水平的 EpCAM 和 CD49f 和低 ERα 表达(图 3 C 和S3 D-S3F),这是腔内祖细胞的特征。L4 组表型显示出高水平的激素受体ERα、孕酮受体 B (PRB) 和AR,以及受体酪氨酸激酶HER2、EGFR和肝细胞生长因子受体(c-MET)(图 3 C 和S3 F)。这些受体与 ERα 或 HER2 的共表达可以赋予抗 ERα 和抗 HER2 治疗的抗性。缺乏 ERα 和 HER2 与抗 ERα 和抗 HER2 治疗的抗性有关。Ki-67 + luminal 肿瘤细胞在所有 luminal 簇组中均发现,并且在 L7 组中最常见(图 S3 G)。

图S3 | 乳腺肿瘤细胞表型的深入分析
此外,L1-L7 组表型在肿瘤亚型中的分布不同。与 HER2 +和 TN 肿瘤相比,L1 组和 L2 组表型指示成熟的管腔细胞和 L4 组和 L5 组强烈表达 ERα 的表型在管腔 A 和 B 肿瘤中更常见(图3F)。增殖组 L7 表型常见于几个 luminal B 肿瘤、几个 luminal A 肿瘤和一个 TN 肿瘤。Luminal B-HER2 +和 HER2 +肿瘤含有来自 L3 和 L6 组的细胞(图3F)。与邻近的非肿瘤组织相比,许多管腔肿瘤含有更少的 K7 +和更多的 K8 +和 K18 +细胞(图 S3H),表明可能由上调的PI3K和 AKT 信号传导诱导的细胞角蛋白转换。
5.表型异常和肿瘤个体性与预后不良的特征
随后,建立了三个计算分数来量化肿瘤异质性的不同方面(图 4A)。表型异常描述了肿瘤细胞表型偏离非肿瘤上皮细胞的程度。肿瘤个体性根据细胞表型量化肿瘤的相似性。肿瘤丰富度表示生态系统内不同共存肿瘤细胞表型的数量。

图4 | 分子表型异常和肿瘤个体性与预后不良的特征
为了描述异常表型,利用自动编码器识别非肿瘤上皮细胞表型并计算每个肿瘤细胞的均方误差 (MSE)(图 S4 A)。结果显示,在上皮簇组 L6、L7、B1 和 B2 中检测到最异常的肿瘤细胞(图4B)。与富含来自 L1 和 L2 组细胞的肿瘤相比,主要包含来自这些簇的细胞的肿瘤更多地偏离近肿瘤组织(图 4 C-4E)。表型异常细胞在高级别肿瘤、大多数 ER -肿瘤、一部分 ER +肿瘤和预后不良的亚型肿瘤中富集(图 4F-4H)。表型异常与缺氧和增殖标志物表达相关(图 4 I),反映了肿瘤生态系统内的异常生长条件。与 4% 的 CA9 -肿瘤细胞相比,约 25% 的 CA9 +肿瘤细胞表现出 EMT 表型。队列中的一些近肿瘤组织样本包含表型异常细胞和高频率的 CA9 +或 Ki-67 +细胞(图 S4 B 和 S4C;表 S5 ),可能代表原位癌前病变导管癌的区域。

图S4 | 肿瘤和非肿瘤组织样本的表型异常和个体性
为了评估肿瘤生态系统的个体性,对所有样本的上皮细胞数据应用了基于图表的方法(图 4 A)。个体性得分表明样本的细胞是否更类似于同一样本的细胞或其他样本的细胞(图 S4 D)。肿瘤显示出比近肿瘤组织更高的个体评分(图 4 J)。重要的是,肿瘤个体性与表型异常相关(图4K),这表明肿瘤细胞与非肿瘤细胞的表型偏离越多,它们在不同患者的肿瘤中发现的可能性就越小(图 S4D)。肿瘤个体性在高级别肿瘤和 luminal B、luminal B-HER2 +或 TN 亚型肿瘤中更为突出(图 S4 E)。ER +肿瘤的个体差异很大,并且与 ERα +细胞的百分比相关(图 4 L 和 4M)。未检测到个体与侵袭淋巴结或远处转移之间的关联(图S4 F)。
为了探索肿瘤丰富度(图 4A),计算了每个样本的每个上皮细胞簇的频率,并报告了 1% 以上的簇数。所有肿瘤和非肿瘤样本都包含来自多个簇的细胞(图 4 N)。值得注意的是,与非肿瘤组织相比,大多数肿瘤的丰富度没有增加,并且肿瘤丰富度与个体性反相关(图4O 和 4P)。在 43% 的肿瘤样本(144 个中的 62 个)中,至少 50% 的肿瘤所有细胞属于一个簇,这可能反映了不同癌细胞克隆的扩增(图 4N)。在 58% 的 3 级肿瘤、33% 的 2 级肿瘤和 35% 的 1 级肿瘤中观察到这种集群优势。在 51% 的 luminal B、50% 的 luminal B-HER2 +和 67% 的 TN 肿瘤中观察到簇优势,但在 luminal A 肿瘤中仅占 29%。在 45 个上皮细胞簇中,37 个簇(82%)占一个或多个肿瘤中所有细胞的至少 50%(图 4 N)。七个优势簇是肿瘤特异性的,四个显示出高表型异常(图 4 B 和S4 G)。
对队列中的 10 个肿瘤进行的分析,尽管新辅助化疗并未显著消退,但揭示了个体表型组成,表明不同的肿瘤细胞表型在治疗中存活(图 4 Q)。这些包括具有 ERα - HER2 -表型和 ERα + HER2 +表型的高度异常肿瘤细胞(图 4B 和 4Q)。从其他四名患者那里收集了同一肿瘤的两个不同区域。在三个案例中,在两个区域都观察到了相似的表型组成。在第四个肿瘤中,优势克隆存在于两个区域,但频率不同,一个区域的增殖特征(5% Ki-67 +)比另一个区域(0.6% Ki-67 +)更具增殖性(图 S4 H)。
6.基于肿瘤生态系统的分类揭示了不同的组和多个肿瘤单例
层次聚类生成的热图显示三组包含许多肿瘤、四组包含三个或四个肿瘤、36 个肿瘤单例和三组非肿瘤样本(图 5)。主成分分析确定了解释不同组之间最高变异性的集群。此外,Tu1-Tu7 组的临床亚型和分级存在异质性(图 S5 A 和 S5B)。

图5 | 肿瘤生态系统中的关系与疾病进展的特征
在 N2 组中发现了所有乳房成形术样本和 54% 的近肿瘤样本,表明生态系统密切相关(图 5A)。与非肿瘤组织相似,在 N2 组中发现的五个肿瘤主要包含 L1 和 L2 组上皮表型、循环 T 细胞 (T16) 和组织驻留巨噬细胞 (M08) (图 5A )。所有 5 个肿瘤均为 luminal A 亚型和低级别,这表明肿瘤在表型上与非肿瘤组织相似,或者这些样本中的肿瘤含量特别低。肿瘤单例通常不共享肿瘤细胞表型,反映了高肿瘤个体性,并且比 Tu1 和 Tu3 组的肿瘤具有更高的表型异常评分和更多的增殖细胞(图 5 C-5E)。Luminal B-HER2 +、HER2 +和 TN 肿瘤要么是小肿瘤组的一部分,要么是单个肿瘤(图 5A)。因此,肿瘤上皮及其免疫环境中的模式都包含肿瘤分层信息。
7.乳腺肿瘤及其免疫环境是相互交织的实体,两者对于分类都很重要
肿瘤细胞和肿瘤-宿主细胞相互作用的网络推动着疾病进展,是药物干预有希望的目标。为了系统地阐明同型和异型肿瘤与免疫细胞的关系,对所有样品中所有细胞表型簇的频率进行了成对 Spearman 相关分析(图 6A -6C;表 S5)。在来自不同簇组的表型之间发现了同型上皮细胞关系(图6A)。非肿瘤管腔表型如 Ep30 和 Ep31(L1 组)是相关的,而肿瘤特异性表型通常分别富集,反映表型优势和肿瘤个体性(图 6A )。L1 和 L2 组以及 Ep39 簇中非肿瘤表型的频率与这些免疫抑制表型呈负相关(图6B ),但与 PD-1 int CTLA-4 - CD38 -表型 T07 和 T18 相关(图 6 B)。肿瘤相关免疫细胞之间的关系分析显示,T-regs 和 PD-L1 + TAM 表型与 PD-1高CTLA-4 + CD38 +的 T 细胞表型相关,表明免疫抑制相互作用(图 6 C)。T-regs 和 PD-L1+ TAM 没有或仅与 PD-1 int CTLA-4 - CD38 - T 细胞表型负相关(图 6 C)。
此外,免疫抑制模式与肿瘤表型异常和个体评分、缺氧和增殖相关(图6D)。免疫抑制性 TAM 和 T 细胞与 ERα +细胞丰度之间的相关性(图 6 D),表明雌激素信号传导是肿瘤生态系统中的塑造力量。

图6 | 乳腺肿瘤及其免疫环境是相互交织的实体
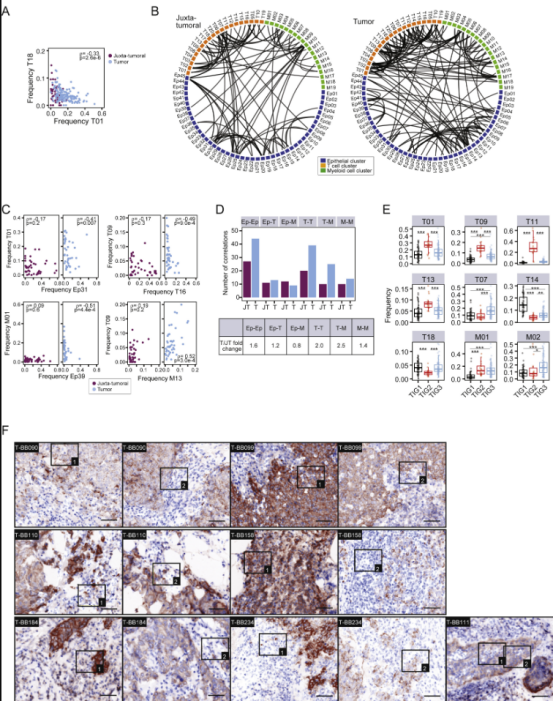
图S6 | 肿瘤生态系统中关系的深入分析
关系分析表明肿瘤免疫环境中有相当大的结构,不同亚型的肿瘤,包括 ER +和 ER -肿瘤,在 TIG2 中分组,提出了 TIG2 中丰富的免疫细胞是否位于肿瘤生态系统近端的问题。因此,除了免疫抑制环境外,TIG2 肿瘤生态系统还包含多个肿瘤细胞群,有可能逃避常见的癌症治疗。
相关讨论
异质肿瘤细胞、浸润性 T 细胞和巨噬细胞之间的交流塑造了乳腺癌生态系统,对疾病进展和临床结果有影响。通过对来自 144 个肿瘤样本、46 个近肿瘤样本和来自四个缩小乳房成形术的组织的 2600 万个细胞进行大规模细胞计数分析,构建了一个广泛的人类乳腺癌生态系统单细胞图谱。该图谱揭示了乳腺上皮细胞和免疫细胞的巨大表型多样性、肿瘤细胞的表型异常和肿瘤个体性,并突出了同型和异型肿瘤-免疫细胞关系,实现了基于生态系统的患者分类。乳腺癌生态系统图谱的建立为识别肿瘤生态系统中临床相关的细胞表型及其关系提供了基础,为将来用于患者分层和精准医学应用提供了理论依据。
现鹿明生物推出质谱流式单细胞蛋白质组技术服务,在推出基于布鲁克tims TOF Pro2 4D蛋白质组质谱平台的单细胞蛋白质组后,再度推出基于质谱流式平台的新一代高通量单细胞蛋白质组技术服务。
鹿明生物引进Starion®星瀚质谱流式系统,具有高通量高分辨率超高速的特点,可实现在单细胞分辨率下同时对数十种目标蛋白的深度分析。进一步丰富鹿明生物单细胞多组学技术服务产品线,为科研工作者提供全面深入的科研技术服务。

猜你还想看
1、Nature | 肠脑轴:16S+代谢+转录揭示间歇性禁食改变肠道菌群,促进神经损伤修复
2、8月“稳” | 恭喜鹿明生物蛋白代谢16篇项目文章发表,总IF:100+
3、Nat Med 新作|Olink蛋白质组学助力化疗免疫治疗一线转移性胰腺癌多组学免疫分析
4、Cell背靠背 | 重大突破!2篇Cell研究人类脑部肿瘤的免疫细胞图谱!
END
空气|撰文
小久|排版
欢迎转发到朋友圈
本文系鹿明生物原创
转载请注明本文转自鹿明生物

我知道你在看哟


点击“阅读原文”了解更多
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章
-
微信文章
-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章
-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章

-
微信文章
-
微信文章

-
微信文章