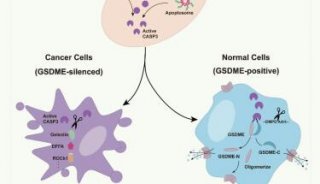
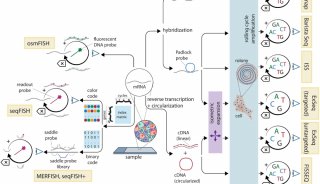
综述:细胞凋亡(三)
细胞凋亡与医学
1.免疫学:
1)胸腺细胞成熟过程中的凋亡:胸腺细胞经过一系列的发育过程而成为各种类型的免疫活性细胞。在这一发展过程中,涉及了一系列的阳性细胞选择和阴性细胞选择过程。以形成CD4+的T淋巴细胞亚型及CD8+的T淋巴细胞亚型;同时,对识别自身抗原的T细胞克隆进行选择性地消除,其细胞克隆死亡的机制主要是通过程序性细胞死亡。因此,正常的免疫系统发育的结局,既形成了有免疫活性的淋巴细胞,又产生了对自身抗原的免疫耐受。耐受机制的形成,主要靠识别自身抗原的T淋巴细胞克隆的程序性细胞死亡机制的活化。
2)活化诱导的细胞死亡:(activation-induced cell
death,AICD)是T淋巴细胞程序性死亡的又一个主要类型。正常的T淋巴细胞在受到入侵的抗原刺激后,T淋巴细胞被激活,并诱导出一系列的免疫应答反应。机体为了防止过高的免疫应答,或防止这种免疫应答无限制地发展下去,便有AICD来控制激活T细胞的寿命。实际上:T淋巴细胞的增殖与T淋巴细胞AICD具有共同的信号通路。T淋巴细胞受到刺激后就开始活化,活化以后的T淋巴细胞如果有生长因子的存在,即发生生殖反应,如果没有或较少的生长因子的存在,则发生AICD。3)淋巴细胞对靶细胞的攻击:免疫活性细胞,特别是淋巴因子激活的杀伤细胞(LAK),是过继性免疫治疗的一种重要形式。在抗肿瘤、抗病毒及免疫调节中具有重要作用。这些免疫活性细胞在攻击肿瘤细胞、病毒感染的细胞时,可诱导靶细胞发生程序性死亡。
2.临床医学:
细胞凋亡之所以成为人们研究的一个热点,在很大程度上决定于细胞凋亡与临床病毒的密切关系。这种关系不仅表现在凋亡及其机制的研究,阐明了一大类免疫病的发病机制,而且由此可以导致疾病新疗法的出现,特别是细胞凋亡与肿瘤及爱滋病之间的密切关系倍受人们重视。
1) HIV病毒感染造成CD4+细胞减少是通过细胞凋亡机制
HIV感染引起爱滋病,其主要的发病机制是HIV感染后特异性地破坏CD4+细胞,使CD4+以及与其相关的免疫功能缺陷,易招致机会性感染及肿瘤,但HIV感染后怎样特异性破坏CD4+细胞呢?近年认为,CD4+T淋巴细胞绝对数显著减少的原因,主要是通过细胞凋亡机制造成的。这不仅阐明了AIDS时CD4+T细胞减少的主要原因,同时也为AIDS的治疗研究指明了一个重要的探索方向。
2)从细胞凋亡角度看,肿瘤的发生是由于凋亡受阻所致
一般认为恶性转化的肿瘤细胞是因为失控生长,过度增殖,从细胞凋亡的角度看则认为是肿瘤的凋亡机制受到抑制不能正常进行细胞死亡清除的结果。肿瘤细胞中有一系列的癌基因和原癌基因被激活,并呈过表达状态。这些基因的激活和肿瘤的发生发展之间有着及为密切的关系。癌基因中一大类属于生长因子家族,也有一大类属于生长因子受体家族,这些基因的激活与表达,直接刺激了肿瘤细胞的生长,这些癌基因及其表达产物也是细胞凋亡的重要调节因子许多种类的癌基因表达以后,即阻断了肿瘤细胞的凋亡过程,使肿瘤细胞数目增加,因此,从细胞凋亡角度来理解肿瘤的发生机制,是由于肿瘤细胞的凋亡机制,肿瘤细胞减少受阻所致。因此,通过细胞凋亡角度和机制来设计对肿瘤的治疗方法就是重建肿瘤细胞的凋亡信号转递系统,即抑制肿瘤细胞的生存基因的表达,激活死亡基因的表达。
3)细胞凋亡的研究将给自身免疫病带来真正的突破
自身免疫病包括一大类难治性的免疫紊乱而造成的疾病,自身反应性T淋巴细胞及产生抗体的B淋巴细胞是引起自身免疫病的主要免疫病理机制,正常情况下,免疫细胞的活化是一个极为复杂的过程。在自身抗原的刺激作用下,识别自身抗原的免疫细胞被活化,从而通过细胞凋亡的机制而得到清除。但如这一机制发生障碍,那么识别自身抗原的免疫活性细胞的清除就会产生障碍。有人观察到在淋巴增生突变小鼠中观察到Fas编码的基因异常,不能翻译正常的Fas跨膜蛋白分子,如Fas异常,由其介导的凋亡机制也同时受阻,便造成淋巴细胞增殖性的自身免疫疾患。
4)神经系统的退行性病变:目前知道老年性痴呆是神经细胞凋亡的加速而产生的。阿尔茨海默病(AD)是一种不可逆的退行性神经疾病,淀粉样前体蛋白(APP)早老蛋白-1(PS1)早老蛋白-2(PS2)的突变导致家族性阿尔茨海默病(FAD)。研究证明PS参与了神经细胞凋亡的调控PS1、PS2的过表达能增强细胞对凋亡信号的敏感性。Bcl-2基因家族两个成员Bcl-xl和Bcl-2参与对细胞凋亡的调节。
线粒体与细胞凋亡
线粒体是真核细胞的重要细胞器,是动物细胞生成ATP的主要地点。线粒体基质的三羧酸循环酶系通过底物脱氢氧化生成NADH。NADH通过线粒体内膜呼吸链氧化。与此同时,导致跨膜质子移位形成跨膜质子梯度和/或跨膜电位。线粒体内膜上的ATP合成酶利用跨膜质子梯度能量合成ATP。合成的ATP通过线粒体内膜ADP/ATP载体与细胞质中ADP交换进入细胞质,参与细胞的各种需能过程。
1951年Glucksmann提出正常脊椎动物发育中的细胞死亡。1966年Saunders提出在形态发生中细胞死亡。1972年Kerr提出细胞凋亡(apoptosis),说明这是在组织动力学方面有广泛作用的一种基本生物学现象。1974年Lockshin提出细胞程序性死亡。美国麻省理工学院Horvitz在研究线虫发育时发现线虫的每个细胞的位置、分裂与命运都是由遗传决定的程序所精确地预先确定的。在构成成虫体时有1090个细胞诞生,131个细胞死亡。1993年袁钧瑛发现线虫的死亡基因ced-3的产物在结构和功能上与哺乳类白细胞介素1β转换酶有同源性。此后,属于同一家族的十几个相关基因陆续在哺乳动物基因组中被发现。统称为胱冬肽酶(caspases)。1994年Hengartner发现线虫的存活基因ced-9的产物与哺乳动物原癌基因bcl-2的产物相似。
细胞凋亡的特征是细胞由于降解酶,主要是水解酶(蛋白酶与核酸酶)的作用,在近乎正常的细胞质膜内趋向死亡。这与坏死时细胞质膜早期破损不同。在细胞凋亡过程中,质膜脂双层丧失二侧不对称性,磷脂酰丝氨酸暴露于细胞表面,从而导致被吞噬。
1 线粒体跨膜电位的耗散与细胞凋亡有密切关系
近年来陆续有报道说明线粒体跨膜电位的耗散早于核酸酶的激活,也早于磷酯酰丝氨酸暴露于细胞表面。而一旦线粒体跨膜电位耗散,细胞就会进入不可逆的凋亡过程。线粒体解联的呼吸链会产生大量活性氧,氧化线粒体内膜上的心磷脂。实验证明,用解偶联剂mClCCP会导致淋巴细胞凋亡。而如果能稳定线粒体跨膜电位就能防止细胞凋亡。
2 线粒体通透性转变孔道
在细胞凋亡过程中线粒体跨膜电位的耗散主要是由于线粒体内膜的通透性转变,这是由于生成了动态的由多个蛋白质组成的位于线粒体内膜与外膜接触位点的通透性转变孔道(PT孔道)(图1)。PT孔道由线粒体各部分的蛋白质与细胞质中蛋白质联合构成。这包括细胞液蛋白:己糖激酶,线粒体外膜蛋白:外周苯并二嗪(benzodiazepine)受体与电压依赖阴离子通道,线粒体膜间间隙蛋白:肌酸激酶,线粒体内膜蛋白:ADP-ATP载体,线粒体基质蛋白:亲环蛋白D(cyclophilin
D)等。凡是能够专一作用于线粒体诱导PT孔道生成的物质,例如苯并二嗪受体的配基原卟啉IX等都能引起细胞凋亡。
3 PT孔道的性质
通过一些实验室的研究,以下诸点值得指出:(1)线粒体内膜通透性转变既是细胞凋亡的必须条件,也是它的充足条件。(2)PT孔道打开后导致线粒体许多功能的致命性变化从而启动了死亡途径。(3)PT孔道作为许多生理效应的感受器(二价阳离子、ATP、ADP、NAD、ΔΨm、pH、巯基与多肽),整合了电生理、氧化还原与细胞代谢状态的信息。(4)PT孔道的组成成分ADP-ATP载体是能量代谢的重要分子,由于ADP-ATP载体是由一个基因家族的几个成员所编码,它的表达有严格的组织专一性。因此,PT孔道在不同细胞中的调节可能稍有不同。(5)PT孔道的作用有自放大的效应。PT诱导ΔΨm耗散,而反过来
mClCCP使ΔΨm去极化会导致PT。一些PT的结果例如ΔΨm 耗散,活性氧的生成本身也会导致PT。这就说明PT会有正反馈,
从而在细胞凋亡中有自摧毁的作用。反过来,如果能防止ΔΨm的耗散,就能避免氧化还原不平衡、磷酯酰丝氨酸的暴露与蛋白酶和核酸酶的激活。
4 PT孔道的开放与关闭
PT孔道有开放与关闭二种构象。PT孔道开放导致细胞凋亡。而PT孔道关闭能防止细胞凋亡。当PT孔道与环孢菌素A(cyclosporin
A)或SH,或米酵菌酸(bongkrek
acid)结合时PT孔道被关闭。在PT孔道开放时线粒体释放细胞凋亡诱导因子(AIF)。AIF可能是一种蛋白水解酶,位于线粒体膜间间隙,它能被蛋白酶抑制剂如N-苄氧羰基-缬氨酰-丙氨酰-门冬氨酰氟甲基酮(N-benzyloxycarbonyl-Val-Ala-Asp-fluoromethylketone)所抑制。此外从线粒体释放的细胞色素C也是一种细胞凋亡诱导因子。虽然苍术苷与米酵菌酸都是ADP-ATP载体的抑制剂,但是它们对PT孔道的作用并不相同。苍术苷促进PT通道开放。这可能与二种抑制剂和ADP-ATP载体的结合部位不同有关。苍术苷只能与ADP-ATP载体的胞液侧结合而米酵菌酸可与ADP-ATP载体的胞液及基质二侧结合。
-
科技前沿

-
科技前沿
-
科技前沿

-
项目成果
-
技术原理

-
技术原理

-
科技前沿
-
技术原理